INTRODUCCIÓN
El propósito de la presente revisión es el somero estudio histórico acerca de las hipótesis que se emitieron para explicar el mecanismo de la transmisión neuromuscular, seguirle la pista a la forma como evolucionaron esas ideas y de que manera el curare jugó un papel determinante en el diseño de las explicaciones que se hicieron del tema. El hecho curioso de que esta droga sea precisamente un antagonista del fenómeno que se trata de describir, se erige sin duda en una curiosa circunstancia científica tanto para la investigación histórica como propiamente médica. Constituye igualmente un desafío, el hecho de intentar la compaginación de esos dos tipos de información, una sobre los acontecimientos y la otra acerca de sus protagonistas que en conjunto y durante más de un siglo discutieron sobre la forma como el impulso central termina por llevar a una respuesta motora y de cómo el curare, en lugar de un obstáculo se desempeñó como un facilitador a los fines de dilucidar la verdad.
1) Teoría Eléctrica
Todo indica que Claude Bernard (1813 – 1878) trabajando con una muestra de curare proporcionada por el químico Théophile Pelouze (1807-1867), eventualmente habría descrito el sitio de acción del veneno unos ochenta años antes del descubrimiento de los primeros neurotransmisores periféricos (Recherches sur le curare. Comptes rendus hebdomadaires de l’Académie des sciences, Paris, 1850, 31: 533-537). En efecto, proponía que el curare actuaba sobre el sistema nervioso motor respetando las propiedades de los músculos lo mismo que a otros elementos histológicos circundantes. También probaba, de algún modo, la independencia fisiológica entre los nervios motores y sensitivos, así como entre los primeros y el músculo. Sin embargo, como el científico se preguntara de qué manera era posible la subsistencia de la contractilidad muscular durante sus experimentos, llegó a imaginar la existencia de la unión neuromuscular pero no en términos químicos. Por el contrario, tenía el concepto de que se trataba de un fenómeno eléctrico o vibratorio que al impedir la transmisión provocaba parálisis. Así planteada, esta propuesta puede considerarse como el punto de partida de lo que llamaremos la teoría eléctrica de la transmisión neuromuscular.
Con demostraciones como aquellas y dada la selectividad del curare, el investigador propuso que este agente podría ser empleado como una herramienta experimental para diferenciar los mecanismos neuromusculares de los musculares propiamente dichos. Pronóstico que se cumplió con el correr del tiempo y prácticamente es la base para el análisis que realizaremos. Por lo tanto, desde el mismo inicio de las consideraciones e hipótesis sobre la neurotransmisión, el curare comienza a jugar un papel preponderante para la explicación del fenómeno.
Bernard, quien apenas ocupó el lugar número 26 entre los 29 estudiantes de medicina de su grupo, emergió de la oscuridad en las sombras de François Magendie, cuando ya tenía 40 años. Se desempeñaba con un mínimo de equipos, todo en un estilo parecido al de su maestro y siendo un escéptico de las estadísticas, nunca predijo la utilidad futura de esta disciplina. Durante mucho tiempo trabajó sin métodos exactos, hasta el extremo de que algunas de sus investigaciones pudieran muy bien ser producto del azar y sus resultados descansar más sobre su brillante personalidad que en estricto apego a principios científicos. Sin embargo entre otros galardones, la Academia de Ciencias le otorgó el gran premio de Fisiología en los años 1849, 1851 y 1853, por sus trabajos sobre la función vasomotora y la actividad del sistema nervioso simpático.
Como hemos apreciado, es cierto que Bernard había sido el primero en demostrar experimentalmente que en una rana curarizada el músculo no perdía su contractilidad al ser estimulado directamente y que si se ligaba previamente una pata, impidiendo el flujo sanguíneo, el nervio no se veía afectado por el curare. Su conclusión fue entonces de que el curare, de alguna manera “paralizaba al nervio”. Pero luego ocurrió que en otro experimento donde sumergió el nervio en una solución de curare, notó que este no perdía su función, concluyendo que el efecto recaía justo en las ramificaciones cerca del músculo. Sin embargo, ocurrió después de un raro experimento adicional, cuando erróneamente concluyó que el curare desacoplaba el nervio de la médula, publicándolo así en su último libro (De la physiologie générale. Paris Hachette, 1872; 230-7).
Fue más bien Edme Félix Alfred Vulpian (5/1/1826 – 18/5/1887) quien refutó esa postrera hipótesis de Bernard y propuso que el curare provocaba parálisis actuando sobre un punto entre el nervio y el músculo. En otras palabras que la acción de la droga recaía sobre lo que se ha llegado a conocer como la placa motora, un concepto que quedó claramente establecido en 1875. Curiosamente, de este trabajo, se hace poca referencia (Leçons sur l’action physiologique des substances médicamenteuses et toxiques. Paris, Doin, 1882, pp. IV, 220, 240, 255, 268) (Figura 1), siendo como son más conocidas de Vulpian sus contribuciones sobre la función vasomotora y el descubrimiento del sistema cromafín en la médula suprarrenal.
Vulpian nació el 5 de enero de 1826 en París y murió de viruela por haber rechazado la vacunación. Su padre, Alphonse André Jean Baptiste, era abogado de la corte y su madre se llamaba Marie Déme Victoire Darnault.

Figura 1. Trabajo de Vulpian menos conocido: Leçons sur l’action physiologique des substances médicamenteuses et toxiques. Paris, Doin, 1882
Ocurrió entonces que para esta época, el recientemente inventado galvanómetro fue aplicado al estudio de algunos problemas físicos y fisiológicos, entre los cuales estaban la actividad nerviosa y muscular. Varios autores entraron en escena después que Nobili, usando el galvanómetro astático que él mismo había diseñado, anunció que el músculo genera corrientes eléctricas durante la contracción. A continuación y entre los nuevos investigadores en electrofisiología se destacó Cario Matteucci (1811-1865), profesor de física en Pisa y uno de los primeros ministros italianos de instrucción pública. Matteucci descubrió que un músculo lesionado, en reposo, se comporta como una fuente de corriente eléctrica, siendo el primer fisiólogo en demostrar que el nervio en una preparación neuromuscular, podía ser estimulado por otro músculo si ambos entran en contacto, sirviendo su propio músculo como un conveniente indicador. Matteucci también notó que cuando se cerraba un circuito eléctrico estimulador, la excitación del nervio se originaba en la región donde se colocaba el cátodo, mientras que cuando se abría entonces era la región del ánodo la excitada. Además, observó que los electrodos de estimulación se polarizaban, es decir que desarrollan un potencial propio después de haber pasado por ellos una corriente eléctrica. Estos descubrimientos desencadenaron un intenso trabajo científico que sin duda sirvió de apoyo a las teorías de Bernard y Vulpian. A partir de la tercera década del siglo XIX, el recientemente inventado galvanómetro fue aplicado al estudio de varios problemas físicos y fisiológicos, entre los que estaban la actividad nerviosa y muscular. Varios investigadores entraron en escena después que Nobili, usando el galvanómetro astático que él mismo diseñó, anunció que el músculo genera corrientes eléctricas durante la contracción. Entre los nuevos actores en esta historia, destacó Emil Dubois-Reymond, quien descubre lo que hoy conocemos como el potencial de acción.
En esos tiempos, de uno u otro modo, se aceptaba que las neuronas no se fusionaban entre sí a ningún nivel de sus ramificaciones. Entonces, lo que seguía a continuación era la pregunta lógica, de como en este contacto entre ellas era posible la transmisión de una onda eléctrica? El sitio no era visible con los microscopios de esa época, de modo que el espacio que las separaba no podía documentarse. De manera que para quienes sustentaban la teoría eléctrica, resultó muy conveniente la aparición de algunos famosos como Emil DuBois-Reymond (Figura No 2), quien incidentalmente también dedicó el grueso de sus investigaciones a los fenómenos bioeléctricos e imaginó los tejidos vivos, entre ellos el músculo, compuesto de “moléculas” eléctricas. Proponía, aunque especulativamente, la existencia de las sinapsis y a propósito diseñó una serie de técnicas e instrumentos para el estudio de los impulsos nerviosos.
DuBois-Reymond nació el 7 de noviembre de 1818, estudió, vivió, trabajo y murió el 26 de diciembre de 1896 en Berlín. Su padre fue un fabricante de relojes en el cantón suizo de Neuchatel y era hermano del famoso matemático Paul DuBois- Reymond. Entró a la Universidad de Berlín en 1836 y fue discípulo de Johanes Müller (1801-1858), quien le obsequió el ensayo de Cario Matteuci sobre los fenómenos eléctricos de los animales. Su tesis de graduación trató sobre los peces eléctricos y fue el sucesor de su maestro hasta su propia desaparición. En 1851 fue admitido en la Academia de Ciencias de Berlín, de la cual llegó a ser su secretario perpetuo a partir de 1867. Por muchos años fue amigo de Helmholtz, también discípulo de Müller.
Du Bois-Reymond era un experimentalista excelente y tenía una gran habilidad para la instrumentación, por lo cual no le tomó mucho tiempo repetir las observaciones de Matteucci sobre la corriente que fluye en un músculo lesionado, y a la cual llamó “Muskelstrom”. Usando la estimulación farádica, determinó que el “Muskelstrom” disminuía cuando se estimulaba el nervio motor, a lo que precisamente llamó una “variación negativa” o “corriente de acción” del músculo. Había usado el término negativo en un sentido algebraico más que eléctrico, para dar a entender la disminución en la amplitud de la corriente registrada. Du Bois-Reymond enunció una ley según la cual la variación en la intensidad del estímulo tiene más influencia sobre la excitación de una fibra nerviosa o muscular que el valor de la intensidad propiamente dicha (ley de Du Bois-Reymond). Igualmente pudo demostrar que un nervio estimulado sufre una variación negativa, similar a la observada en el músculo y que en realidad constituye el impulso nervioso.

Figura 2. Emil DuBois-Reymond, retrato y dibujo original de uno de sus experimentos.
Du Bois-Reymond fue aún más lejos. Estudiando nervios aislados con su sensible galvanómetro, descubrió en 1845 la existencia de cambios electromotrices en el tejido nervioso, similares a los descritos en el músculo. Observó que si los electrodos del galvanómetro eran aplicados sobre la superficie del nervio, alejados de los extremos cortados, esta superficie era equipotencial. Sin embargo, si uno de los electrodos era colocado cerca del extremo cortado mientras que el otro estaba alejado, el electrodo cercano al extremo cortado era negativo con respecto al segundo. También encontró que cualquier lesión producida por calor o compresión sobre una región previamente intacta del nervio la convertía en negativa con respecto a las regiones aún no lesionadas. Por lo tanto y en vista de que estas corrientes eran registradas en nervios en ausencia de estimulación, fueron llamadas por él como corrientes de reposo.
En otra faceta del tema que estamos tratando, la idea de que alguna sustancia química actuara como un mecanismo disparador de una respuesta fisiológica, también había sido sugerida por Dubois-Raymond. En efecto, propuso que ciertos químicos como el amonio y el ácido láctico podían estar envueltos en la activación de la contracción muscular a través de los nervios. Pero esto no fue más que una gran pieza de razonamiento intuitivo, pues se basaba principalmente en el incremento que se observa en la concentración de esos compuestos en la sangre después de un ejercicio muscular importante y que tiene lugar en forma natural.
Uno de los colaboradores de Du Bois-Reymond fue el fisiólogo y físico.
Hermann Ludwig Ferdinand von Helmholtz quien había nacido en 1821, en Potsdam y era el hijo mayor de un maestro de literatura y filología en la escuela secundaria de la ciudad. Helmholtz más bien sentía inclinación por la física, pero la precaria situación económica familiar le hizo aceptar una beca para estudiar medicina en el Konigliches Medizinisch- Chirurgisches Friedrich-Wilhelm Institut en Berlín en 1838. Esta beca le significaba el compromiso de trabajar como médico militar, lo cual hizo desde 1843 hasta 1848. Durante su estadía en el instituto de medicina, Helmholtz se convirtió en uno de los más aventajados estudiantes de Johannes Müller, quien paradójicamente no creía en la posibilidad de los resultados que se obtuvieron en el área de investigación de su alumno. A pesar de lo cual describieron juntos el origen de las fibras nerviosas que se derivan a partir de las células ganglionares.
En 1848 Helmholtz comenzó a dar clases en la Academia de Artes de Berlín y un año más tarde fue nombrado Profesor de Fisiología y Anatomía Patológica en la Universidad de Konigsberg. Fue aquí donde realizó su famoso experimento sobre la velocidad del impulso nervioso. En 1858 se cambió a Bonn, en 1868 a Heidelberg y en 1871 ocupó la cátedra de Física en Berlín. En 1882 el Kaiser Wilhem I lo elevó a la nobleza, con lo cual su nombre cambió a von Helmholtz. Fue además Presidente de la Sociedad de Física de Berlín, lugar en el que permaneció hasta su muerte en 1894, a la edad de 73 años.
La breve comunicación de dos páginas en la cual Helmholtz reporta la velocidad de conducción nerviosa en 1850 y que estima en 27,25 m/s, puede ser considerada como uno de los puntos de partida de la neurofisiología y biofísica modernas y cuya versión completa publicó dos años más tarde.
a) La sinapsis
Pero el nombre sinapsis propiamente dicho (del griego que puede ser traducido como: enlace y que se refiere al proceso de comunicación entre neuronas) en realidad fue acuñado por Charles Scott Sherrington, mientras redactaba un capítulo relacionado con el sistema nervioso central para un libro de texto editado por Foster en 1897. La palabra sinapsis viene de sinapteína, que Sir Sherrington formara con los vocablos griegos sin, (juntos), y hapteina, (con firmeza). Correspondientemente el término “neurona” habría de ser introducido por W. Waldeyer en 1891, pero sin referirse a las conexiones intercelulares.
Para esta época lucía muy atractivo imaginarse una sinapsis eléctrica, incluso hasta con pequeñas chispas atravesando el espacio entre las neuronas. Dicho de otro modo, que el papel de los nervios en ese caso pudiera ser similar al de un cable eléctrico conduciendo pasivamente los mensajes que se originaban en el cerebro. Este concepto había sido poco cuestionado ya que, por otra parte, estaba en concierto con las enseñanzas de Descartes, quien creía que los músculos respondían a unos espíritus llevados por los nervios. Como ya hemos descrito, debe añadirse que posteriormente, con el advenimiento de la electricidad y de los métodos para detectarla (galvanómetros), fue posible individualizar los nervios motores y hasta producir respuestas musculares después de aplicaciones eléctricas.
Entre las críticas que en este momento se le formularon a la teoría eléctrica, llamaba la atención el hecho de que, en los reflejos propioceptivos existiera un pequeño retardo. Este era precisamente, el caso con el tendinoso, reflejo espinal que solo tiene una sinapsis entre las neuronas sensitivas y motoras; por lo tanto, si la transmisión fuera eléctrica ese retardo no existiría. En otro caso, como el corazón, los sistemas simpático y parasimpático mostraban acciones diferentes a nivel de su sinapsis, una excitatoria y otra inhibitoria, lo cual sería difícil de compaginar con una transmisión eléctrica, ya que se trata del mismo elemento possináptico. En cuanto a un tercer tipo, como la transmisión neuromuscular, también se apreció un mínimo retardo que transcurría entre la llegada del impulso eléctrico a las terminaciones del nervio y el comienzo de la contracción muscular, pero con simplicidad fue atribuido al tránsito a través de la hendidura sináptica.
En los trabajos de Sherrington (Figura 3), se resaltan un buen número de propiedades de los arcos reflejos a nivel espinal y de cómo son influenciados y modulados por estructuras en el cerebro y cerebelo, entre otras. De igual modo describía este autor la interrelación entre la excitación e inhibición central como fundamentos de una integración, concepto que plasma en su famosa obra “La acción integrada del Sistema Nervioso”, publicada por primera vez en 1906. Sherrington explicaba enteramente estos fenómenos que estamos comentando en términos de cambios eléctricos a través de la pared membranosa de las células nerviosas involucradas, reforzando de manera decisiva la teoría eléctrica.
Sherrington nació el 27 de noviembre de 1857, en Islington, Londres. Durante su infancia murió su padre y su madre se casó de nuevo. Comenzó sus estudios de medicina en el Saint Thomas Hospital y posteriormente superó con éxito el examen del Colegio Real de Cirujanos. Tras una breve estancia en Edimburgo, se marchó a Cambridge en 1879, donde realizó la mayor parte de su carrera médica. En 1913 fue nombrado catedrático de Fisiología de Oxford, puesto que desempeñó hasta 1936. Durante su estancia en Oxford también publicó su conocido libro “Fisiología de los Mamíferos: Un curso de ejercicios prácticos”, que utilizaba para la enseñanza. En 1922 le fue concedida la Cruz del Gran Cordón de la Orden del Imperio Británico y en 1924 la Orden del Mérito. En 1932 se hizo acreedor al Premio Nobel, compartiéndolo con Edgar Douglas Adrian. Murió en Eastbourne, víctima de un fallo cardíaco en 1952.
Pero estos investigadores ni sus experimentos pudieron identificar el verdadero mecanismo íntimo de los resultados que observaban y fue así que la llegada del siglo XX coincidió con los trabajos de otros fisiólogos de Cambridge como Walter Hollbrook Gaskell quien estudiaba un sistema de nervios que parecían, tanto anatómica como funcionalmente distintos del sistema nervioso central y que más tarde en 1916 llamó “sistema nervioso involuntario”.
Gaskell, quien había nacido el 1 de noviembre de 1847 en Nápoles y muerto el 7 de septiembre de 1914 en Great Shelford, había recibido su educación en el Trinity College, Cambridge. Fue elegido como miembro de la Real Sociedad en 1882 y ganador de la Medalla Real en 1889.
Aquellas ramas nerviosas que tanto llamaron la atención de Gaskell, ya se conocían desde que en 1847, Charles E. Brown-Sequard (1817-1894) observó que su estimulación causaba enrojecimiento de la cara. Este investigador había sido discípulo de Bernard al cual sucedió en su puesto del Collège de France. Nació en Puerto Luis (Isla Mauricio) en 1817 y era hijo de un inglés y de una francesa. Se trasladó a París para estudiar medicina y recibió el grado de doctor en 1845. En 1856 obtuvo el premio de Fisiología experimental y, entre 1858 y 1865, publicó la revista Journal de Physiologie de l’homme et des animaux. En 1863 fue nombrado profesor de Fisiología en Harvard, pero más tarde, tras la muerte de Claude Bernard, ocupó el puesto de este y obtuvo la ciudadanía francesa.

Figura 3. Sherrington: retrato y portada de una versión de su obra más conocida.

b) El capítulo Langley
John Newport Langley, por su parte, pensaba que el mencionado sistema involuntario estaba realmente compuesto por dos secciones separadas que llamó simpático y parasimpático y que al actuar concertadamente forman una red primitiva pero eficiente de retroalimentación. Langley reconoció que era difícil de explicar de qué manera unos efectos opuestos se podían producir por dos sistemas de nervios aparentemente idénticos, si de acuerdo a la idea convencional, los nervios actuaban simplemente llevando la misma forma de “corriente eléctrica” y por lo tanto deberían provocar el mismo tipo de respuesta en el órgano que inervan. Entonces, para explicar que tales fenómenos eran posibles, debían cada uno utilizar un código diferente para transmitir su señal, por lo cual para resolver el problema era necesario conseguir un mecanismo mediante el cual este código especial pudiera trabajar. Fue cuando se le ocurrió la idea de una “llave” química específica para cada uno de ellos, llegando tan lejos en sus deducciones como para proponer que dos sustancias químicas diferentes se liberaban en las terminales del nervio, una para cada sistema y de allí que pudieran observarse efectos diferentes.
Langley había nacido en Newburry el 10 de noviembre de 1852. Hijo de un maestro de escuela privada, fue educado inicialmente en su hogar y luego en la Escuela Exeter antes de ingresar en el Colegio de San Juan de Cambridge en octubre de 1871. Con el propósito de hacer carrera en el Servicio Civil estudió matemáticas, historia y otras materias literarias, pero cambio sus planes en el segundo año cuando comenzó ciencias naturales. Aún antes de graduarse, su profesor Michael Foster del Colegio de la Trinidad lo involucró en sus investigaciones sobre Pilocarpina, un alcaloide que había aislado de las raíces de jaborandi (Preliminary notice of experiments on the physiological action of jaborandi: Br Med J 1875;1:241-242). En 1878 recibió su título, después de un año de estar desempeñándose como asociado. Estuvo entre los primeros investigadores experimentales que utilizaron la anestesia con sus animales y así lo hizo constar explícitamente en sus publicaciones. Algunos de sus trabajos sobre la secreción salival fueron realizados en el laboratorio de Wilhelm Kühne (1837 – 1900) en Heidelberg donde pasó algún tiempo en 1877. En 1883 fue elegido Asociado de la Real Sociedad de Londres y al año siguiente profesor de histología. Fue durante estos años cuando se interesó en la estructura y funcionamiento del sistema nervioso vegetativo o involuntario que había sido descrito por Gaskell. En 1886 fue capaz de distinguir las fibras viscerales (simpático) con origen torácico de las provenientes de la región cervico-cranial y sacro (parasimpático).

Figura 4. J.N. Langley: retrato y título original de su trabajo publicado en el J Physiol December 30, 1905 33:374.

Para 1894, Langley (Figura 4) se hizo editor del Journal of Physiology en el cual publicó su primer trabajo sobre las “sustancias receptivas” (1905). En 1899 fue nombrado Presidente de la Asociación Británica para el Avance de las Ciencias. Mostró gran talento para el deporte del patinaje sobre el hielo, para el cual diseño algunos reglamentos. Murió el 5 de noviembre de 1925 en Cambridge, donde vivía desde su matrimonio con Vera Kethleen Forsythe-Grant en 1902.
En su momento, Langley se inspiró en los trabajos de George Oliver y Edgard A Schäfer, quienes al inyectar un extracto de médula adrenal a perros habían notado una marcada elevación de la presión sanguínea (The physiological effects of extracts of suprarenal capsules. J Physiol (Lond) 1895; 18: 230 – 280). Después de repetir este experimento, Langley concluyó que dichas sustancias actuaban en la fibra nerviosa en lugar de hacerlo sobre la muscular, añadiendo que estos agentes “…no estimulan directamente la sustancia contráctil sino que lo hacen en una porción protoplasmática accesoria...”.
En compañía de William Lee Dickinson (1863-1904) habían descrito el gran potencial de la nicotina como herramienta de investigación y juntos descubrieron que bloquea la conducción nerviosa en los ganglios simpáticos, mientras que la muscarina y su antagonista atropina lo hacen sobre las terminaciones periféricas del vago que se dirigían al corazón, conceptos que corrigió después de las descripciones de Ramón y Cajal.
Luego el investigador dirigió su atención al efecto de la nicotina sobre el músculo y el antagonismo recíproco con el curare. Fue durante estos experimentos cuando escribió: “…Ni los venenos ni el impulso nervioso actúan directamente sobre la materia contráctil del músculo, más bien lo hacen sobre una sustancia accesoria, donde se recibe el estímulo que posteriormente es transferido a los elementos contráctiles y que podríamos llamarla sustancia receptora del músculo…”. Estos conceptos los hizo extensivos a la manera de actuar de otros alcaloides como la pilocarpina, atropina y estricnina así como también de algunas secreciones internas como las hormonas tiroideas y la secretina. En esta etapa, esas fueron las mayores contribuciones al desarrollo de la investigación en fisiología neuromuscular y de la transmisión nerviosa y en la cual también tuvo participación el curare.
Todos esos términos como “porción protoplasmática accesoria”, “sustancia accesoria” y “sustancia receptora”, son difíciles de considerar sin asociarlos con el concepto de “receptores” proveniente de la idea originalmente expuesta por Paul Ehrlich (1854 – 1915), sobre la base de la teoría de la “cadena lateral” de sus investigaciones inmunológicas. En estas, Ehrlich ciertamente, las describe pero sin mencionar el nombre propio, cuando especula que aquellas substancias receptoras bien pudieran ser una cadena lateral de moléculas receptoras. Langley, quien siempre se cuidó de no utilizar el término “receptor”, enfatizó que había llegado a sus conclusiones por un camino diferente al de Ehrlich y cuando este aún no había expuesto su teoría
Las observaciones de Langley sobre el sistema nervioso autónomo lo llevaron a concluir que este era responsable de las funciones subconscientes que mantenían la normalidad del cuerpo, tal como de manera elemental fuera previsto por Claude Bernard cincuenta años antes cuando se expresó sobre el control que debía existir “...para mantener la constancia del medio interno...”, así como también fue la base para lo que más tarde Walter Bradford Cannon (19/10/1871 – 19/10/1945), un fisiólogo de Harvard denominó homeostasis (“The Wisdom of the body”: 1932) y quien junto con el fisiólogo mexicano Arturo Rosenbleuth, publicó la obra “Sistemas Autónomos Neuro-Efectores” donde sintetizan los conceptos de la “mediación química”.
promover sus teorías empezando por exponerlas en la Croonian Lecture to the Royal Society (Langley JN: Croonian Lecture 1906: On nerve endings and on special excitable substances in cells’. Proc R Soc Lond Series B, 1906; 78: 170 – 194), siguiendo a Viena y luego en el Séptimo Congreso Internacional de Fisiólogos en Heidelberg, donde tuvo que confrontar un trabajo crítico de un antiguo discípulo Rudolph Magnus (1873 – 1927) sobre el mutuo antagonismo entre nicotina y curare en preparaciones desnervadas y donde este argumentaba que si bien las observaciones de Langley eran acertadas, en cambio no aportaban indicios sobre la localización del efecto. Estas divergencias se prolongaron en el tiempo y Langley posteriormente propuso, en primer lugar, que al provocarse la denervación también resultaban dañadas las sustancias receptoras y que a todo evento debían existir sustancias receptivas diferentes de acuerdo al veneno utilizado (nicotina o curare, en este caso).
Después se enfrentó al reto de la teoría expuesta en 1908 por Walter Straub (1874 – 1944), para quien se trataba de un conflicto de concentraciones y gradientes para explicar el mecanismo de acción de las drogas. Straub trabajaba con un molusco (Aplysia) y proponía que la muscarina actuaba en el corazón mientras se encontraba dentro de la célula, condiciones que se mantenían en tanto lo hacía la concentración exterior, pero ningún incremento podía modificar en ese momento la cantidad dentro de la célula (Zur kinetic der makirinwirkung und des antagonismos muskarin-atropin: Pflüger’s Archiv für gesamte physiologie 1907; 119: 127-151). Langley entonces replicó que mientras el veneno permaneciera unido a la sustancia receptora se originaban estímulos en dirección a la célula, pero que si se saturaba no podía dar lugar a nuevos estímulos e igualmente hizo extensiva esta explicación al caso del antagonismo atropina-muscarina, durante el cual, cuando la primera se combina con la sustancia receptora impide la acción de la segunda.
Langley igualmente se vio obligado confrontar a Walter Ernest Dixon también de Cambridge, quien había demostrado que la estricnina no se combinaba químicamente con el tejido nervioso y supuestamente esto probaba que tampoco se unía a ninguna sustancia receptora. Aseguraba Dixon textualmente que: “...cuando un músculo se contrae o una glándula secreta o una terminación nerviosa es excitada, la causa en cada uno de estos casos puede deberse a la liberación de alguna sustancia química, no necesariamente libre en la circulación, como la secretina, pero más probablemente liberada en un lugar sobre el cual debe actuar...“ (On the mode of action of drugs; Med. Megaz. (Lond). 1907;16:454-457).
Pero Langley finalmente consiguió apoyo a su teoría por parte de un discípulo: Archibald Vivian Hill (posteriormente ganador un Premio Nobel en 1922) quien analizó matemáticamente el experimento de Langley y confirmó la posibilidad de una combinación química con algún componente del tejido muscular. Hill había nacido en Bristol el 26 de septiembre de 1886 y su primera educación la recibió en la Escuela de Blundell, Tiverton, donde obtuvo una beca para el Colegio de la Trinidad en Cambridge con el objeto de estudiar matemáticas, pero después de graduarse su profesor Walter Morley Fletcher, lo impulso a dedicarse a la fisiología.
2) La Etapa Intermedia
En 1900, Dixon había reportado sobre un transmisor nervioso químicamente parecido a la muscarina. Se trataba de un veneno producido por un hongo que actuaba en el sistema nervioso parasimpático. Aparentemente esto no lucía como un conocimiento nuevo, pues algunos aseguran que ya para 1869, Schmiedeberg y Koppe conocían del efecto depresor cardíaco de la muscarina (Das muscarin. Das giftige alkaloid des fiegenpilzes. Vogel, Leipzig). El envenenamiento por esta sustancia se asemeja mucho a la estimulación del nervio Vago, caracterizándose por un enlentecimiento de la frecuencia cardíaca y un aumento de la actividad intestinal que también pueden acompañarse de salivación profusa y constricción de la pupila. No tardó entonces en proponerse que la activación del Vago pudiera deberse a la liberación de pequeñas cantidades de muscarina
Otra evidencia en el mismo sentido se debe a: T.R. Elliot, quien basado en los resultados de Langley, inyectó adrenalina a un perro y observó una respuesta similar a la provocada por la estimulación eléctrica de sus nervios simpáticos. En el resumen que presentó ante la Sociedad Británica de Fisiológica en 1904 (Elliot TR: On the action of adrenalin. J Physiol (Lond) 1904; 31: 20 – 21) hizo la brillante sugerencia de que la epinefrina liberada de los nervios
simpáticos induce las respuestas fisiológicas de los órganos que son inervados por aquellos. Puesto en sus propias palabras, Elliot se expresó del siguiente modo: “…El punto en el cual se recibe la excitación de un estímulo químico y este se transforma en lo que pueda ser la causa o el cambio de tensión del músculo, es posiblemente un mecanismo desarrollado por la célula muscular…”. Langley a quien no le gustaban las teorías estuvo persuadiendo a Elliot para que no publicara sus ideas hasta obtener un mayor número de pruebas y de hecho el concepto de neurotransmisión química no fue mencionado en su trabajo inicial. Pero Elliot todavía hizo una segunda publicación en el mismo sentido, confirmando sus presunciones preliminares (J Physiol (Lond) 1905;32:401-467).
El siguiente gran paso en la ruta hacia el esclarecimiento de la transmisión nerviosa, se puede atribuir a Reid Hunt quien observa como los extractos suprarrenales libres de adrenalina son más bien capaces de bajar la presión sanguínea y responsabiliza a la colina por dicho efecto. Para que más tarde junto a Rene de M. Taveau identificara a la acetilcolina como un agente mucho más potente que la colina para bajar la presión arterial (On the physiological action of certain choline derivates and new methods for detecting choline. Br Med J 1906;2:1760).
De ninguna manera puede considerarse como menor el hecho de que en 1902, Ernest Henry Starling (1866-1927), y William Maddock Bayliss (1860-1924) habrían realizado su mayor aporte a la medicina: el descubrimiento de la secretina, la primera hormona, nombre este derivado de una palabra griega que puede traducirse como excitación o despertar. Durante el curso del experimento que ambos realizaron para demostrar el hallazgo, perforaron el intestino de un perro anestesiado, en el cual se vio cómo, al mezclarse el ácido clorhídrico con los restos de comida, inmediatamente se formaba una sustancia química en las células epiteliales del duodeno, a la que llamaron secretina. Esta sustancia, a través de la circulación de la sangre, entraba en contacto con el páncreas, el cual, a su vez, estimulaba la secreción de los jugos digestivos al intestino a través del conducto pancreático. Fue de este modo como tuvo lugar la presentación de la teoría del control hormonal de las secreciones internas.
Sir Bayliss nació en Butcroft, Wednesbury el 2 de mayo de 1860 y falleció en 1924, hijo único de Moses Bayliss, un fabricante de tornillos y de Jane Maddock. Fue al Colegio Universitario de Londres en 1881 y en 1885 al Colegio Wadham de Oxford, obteniendo la más alta calificación cuando terminó la Escuela de Ciencias Naturales y Fisiología. En 1915 publicó su clásico libro: “Principios de Fisiología General” y de la cual en vida aparecieron cuatro ediciones. Bayliss, se había casado en 1893 con Gertrude, la hermana de Starling y quien, por lo tanto era su cuñado. Incidentalmente también, sus personalidades fueron complementarias; pues Starling era apresurado, incauto, impaciente y agresivo, mientras que Bayliss fue cauteloso, metódico y paciente.
Por su parte, Starling nació en Londres en 1866. Su padre era abogado, trabajaba en Bombay y regresaba a Inglaterra cada tres años. Su madre, Ellen Watkins, residía en Gran Bretaña y cuidaba de los hijos, de los cuales Ernest era el mayor. Se formó en Islington (1872-1879) y en la Escuela del Colegio Real (1880-1882). Comenzó sus estudios de medicina en la Escuela de Medicina del Hospital Guy en 1882 y se graduó en 1889. En 1885 pasó un verano en el laboratorio de Willy Kühne en Heilderberg. En 1889 fue “demonstrator” de Fisiología en el Hospital Guy y en 1890 empezó a trabajar también en el laboratorio de Sir Edward Albert Sharpey-Schäfer (1850-1935) en el University College. Allí comenzó una fructífera relación con William Maddock Bayliss y su primer artículo conjunto apareció en 1891. Murió en un barco durante una travesía por el Caribe el 2 de mayo de1927 y fue enterrado en Kingston, Jamaica.
Los años que comprenden la década de 1930 fueron de mucha especulación para sostener la teoría eléctrica en el proceso de la transmisión sináptica central, ya que como parte de su final dramático se había descrito en numerosas especies inferiores, tanto invertebrados como vertebrados.
a) El capítulo Eccles
Cronológica, científica y hasta “geográficamente” proponemos reconocer en este momento, que hemos arribado al punto de cruce, cuando se pasa de la concepción eléctrica a la química de la neurotransmisión y la escena será copada durante muchos años por un personaje clave que fue la figura relevante en los dos actos de esta obra: John Carew Eccles. También consideramos que la descripción somera de sus conceptos sobre la teoría eléctrica son los que relatan del mejor modo científico su contenido.
En 1938, Eccles y W. J. O’Connor pudieron por primera vez registrar potenciales eléctricos de la placa final en la superficie de fibras musculares. La creencia de que esos fenómenos eran precedidos por potenciales de acción musculares conjuntamente con la acción que ellos adjudicaron a la fisostigmina y al curare, llevó a Eccles a negar el papel de la acetilcolina en la transmisión neuromuscular y a proponer una opción que llamó “la teoría del detonador”. Como también había podido registrar otras respuestas y estos nuevos resultados le parecieron consistentes con la hipótesis eléctrica, así lo publicó en Nature de diciembre de 1945 y lo presentó en la Academia de Ciencias de Nueva York en febrero de 1946 durante su primera visita a Estados Unidos. Este mismo año recogió los eventos eléctricos extracelulares asociados con la excitación monosináptica, los cuales aparentemente también concordaban con las predicciones de la teoría eléctrica, por lo tanto propuso que los potenciales sinápticos producían corrientes que fluían a través de los axones terminales y pasivamente actuaban sobre la excitabilidad de las motoneuronas. Adicionalmente, había descubierto que no se observaban cambios en la transmisión sináptica si la preparación permanecía sumergida por largo tiempo en una solución de acetilcolina y esto lo hizo considerar que la transmisión química jugaba un papel subordinado y despreciable. Era posible, igualmente, que la aparente prolongación de ciertos efectos producidos por los anticolinesterásicos y por el curare pudiera atribuirse a una acción simultánea con la acetilcolina y de esa manera se llegaban a provocar corrientes de acción como impulsos presinápticos. Entonces es notorio, una vez más, que la existencia y utilización del curare también fue la herramienta de investigación que mejor sirvió al diseño de lo que fue la versión más importante de la teoría eléctrica de la neurotransmisión.
Pero no cabe duda que esta atractiva hipótesis fuera a tropezar con serios escollos. El flujo de información en la cadena neural es unidireccional, siempre en el sentido axón-dendrita, de manera que si la sinapsis fuera eléctrica, como proponían, sería difícil imaginar la manera de impedir el flujo en dirección opuesta en caso de excitarse algún elemento possináptico. Pero este obstáculo pareció resuelto en el momento cuando se hizo evidente la existencia de sinapsis excitatorias y sinapsis inhibitorias. En segundo lugar, aunque el potencial de acción tiene siempre la misma polaridad, todavía sería difícil aceptar que una sinapsis puramente eléctrica pudiera servir al mismo tiempo para la excitación o la inhibición y en tercer lugar, obraba en contra de la teoría eléctrica, que algunos reflejos monosinápticos muestran un ligero retardo que no existiría si la transmisión fuese eléctrica.
Eccles nació el 27 de enero de 1903 en Norticote, un suburbio de Melbourne, el primero de dos hijos de un par de maestros de escuela en Victoria donde ambos a su vez también habían nacido. A los 12 años empezó la secundaria y cuatro años más tarde entró a la Universidad de Melbourne después de haber estudiado matemáticas. Fue el primero de su clase y del Estado, haciéndose acreedor a una beca universitaria. A los 17 años inició sus estudios de medicina y en razón al éxito demostrado tanto académico como deportivo, ganó varios premios entre las universidades australianas. En 1925 se graduó de bachiller en medicina y cirugía como el primero de su clase y después de seis meses como residente, abandonó la ciudad con destino a Oxford.
Dos lecturas habían impactado a Eccles hasta este momento de su vida: el “Origen de las Especies” de Darwin, cuando estudiaba Zoología en primer año y el libro de Sherrington: “Acción integrada del sistema nervioso”. Fue aceptado en el Colegio de la Magdalena, donde Sherrington precisamente estaba asociado. Como le fuera presentado inmediatamente dio comienzo a una profunda y cercana amistad que persistió hasta su muerte. Se graduó con honores en las escuelas de fisiología y bioquímica y esto le valió una nueva beca a mediados de 1927, esta vez para el Exeter College donde comenzó a estudiar para su PhD siempre bajo la supervisión de Sherrington y en condición de Asociado Menor de Investigación.
En su vida personal, Eccles contrajo matrimonio en 1928 con Irene Millar, oriunda de Motuela, Nueva Zelanda, con quien procreó nueve hijos. Después de cuarenta años se divorciaron y se casó con Helena Taborikova, una investigadora nacida en Checoslovaquia.
En 1934 logró una posición permanente en Oxford, pero el reemplazo de Sherrington en 1935 le produjo una gran desilusión y aunado a la creciente situación de incertidumbre política en Europa, lo indujeron aceptar la dirección del Instituto Kanematsu de Patología en el hospital de Sydney. Aquí continuó sus investigaciones sobre la transmisión de los impulsos del nervio al músculo, esta vez acompañado por Bernard Katz hasta 1943.
Entre tanto sus relaciones con la junta del hospital se hicieron pocos armoniosas, dados el escaso entendimiento que mostraban por las investigaciones básicas que realizaba y su relevancia clínica, lo cual llevó a Eccles a presentar su renuncia en octubre de 1943 y aceptar el cargo como Jefe de la Cátedra de Fisiología en la Escuela de Medicina de la Universidad de Otago en Dunedin, Nueva Zelanda. Durante la IIa Guerra Mundial sirvió en el ejército australiano como consejero en diversos comités de especialidades médicas y en la síntesis de suero sanguíneo. Preocupado por la pesada carga docente que limitaba su competitividad en las investigaciones neurofisiológicas, aceptó la Cátedra de Fisiología en la Escuela Médica de Investigación John Curtin de la Universidad Nacional Australiana en Canberra, para donde se mudó en septiembre de 1952. Hasta que en 1966 se vio forzado a retirarse por haberse cumplido el período reglamentario de edad.
Fue entonces cuando aceptó incorporarse como miembro del Instituto de Investigaciones Biomédicas que recientemente había creado la Asociación Médica Americana en Chicago. Eccles mismo describe este período como “…el más breve, el menos exitoso y más infeliz de mi carrera como investigador…”. Aunque en esa etapa había continuado su estudio de la corteza cerebelar, pensó que a su edad no podría continuar recibiendo apoyo financiero para sus trabajos, lo cual sumado a desavenencias con su grupo lo inclinó por aceptar otra opción. En 1968 esta nueva oportunidad se le ofreció como Profesor Distinguido de Fisiología y Medicina y Asociado de Investigación Henry C y Bertha H. Buswel en la Escuela de Medicina de la Universidad del Estado de Nueva York en Buffalo.
En 1975, Eccles se retiró voluntariamente y se mudó a la ciudad de Contra en el condado suizo de Ticino, que describió como: “…una montaña idílica para dedicarse a trabajar sobre el problema mentecerebro…” y donde murió el 2 de mayo de 1997.
En 1930 se interesó en la naturaleza de la transmisión sináptica, particularmente en la cuestión fundamental de cómo las señales eran transferidas de una célula nerviosa a otra. En 1931 llevó a cabo un estudio con G. L. Brown que aparentemente lo introdujo en la creencia de que un proceso eléctrico pudiera ser el responsable de las características de duración en la transmisión sináptica
Pero aconteció que desde 1949, Eccles (Figura 5) se encontraba revisando su hipótesis eléctrica a través de una modificación sobre la técnica que empleaba y en vista de los efectos inducidos por algunos inhibidores anticolinesterásicos, pareció convencerse de que la transmisión a nivel de la unión neuromuscular podría ser un proceso químico y mediado por la acetilcolina. Los registros efectuados el 20 de agosto de 1951 mostraron un resultado contrario a las predicciones de su anterior teoría y reconoció la mediación de transmisores químicos. Aunque al principio solo aceptó el cambio para la excitación monosináptica, los trabajos sobre la regeneración de los nervios finalmente le sugirieron la secreción de sustancias específicas en las motoneuronas. A propósito de lo cual escribió textualmente: “…el bloqueo sináptico producido por el curare en el músculo esquelético, actúa como un depresor específico de las respuestas provocadas en la placa motora por la acetilcolina, proporcionando una explicación obvia al bloqueo sináptico que produce…”. Fue de esta manera como una vez más el empleo del curare fue la clave para el desarrollo de la nueva hipótesis química.
Mientras duró su estadía en Canberra, Eccles realizó trabajos sobre los cambios químicos que tienen lugar en la sinapsis. Demostró que diferentes células nerviosas producen la liberación de una sustancia, probablemente acetilcolina y esta a su vez provoca el paso de sodio y potasio alterando la polaridad de la carga eléctrica, constituyendo entonces la forma definitiva como los impulsos se comunican o inhiben a otras células nerviosas. Estos resultados junto a los hallazgos de Alan Hodgkin y Andrew Huxley, sus estrechos colaboradores, los hizo merecedores del Premio Nobel 1963 en Fisiología y Medicina.

Figura 5. J. C. Eccles: Retrato y primera página de uno de sus más importantes trabajos sobre la transmisión eléctrica.

3) Desarrollo de la Teoría Química
a) El capítulo Dale
El prolongado debate entre los promotores de las teorías eléctrica y química de la transmisión sináptica también se había extendido por las décadas de 1940 y 1950, siendo uno de los más significativos en la historia de la neurociencia del siglo XX. Consideramos que para esta nueva etapa, el mejor punto de partida debe ser Sir Henry Hallet Dale, cuyos primeros trabajos asociado con Arthur Ewins versaron sobre los efectos fisiológicos de la ergotamina, incluyendo la identificación precisa de su alcaloide activo y la reversión sobre la acción de la adrenalina y de los nervios simpáticos. Como resultado de otras investigaciones había propuesto que la adrenalina solo reproducía parcialmente los efectos simpáticos y para 1910 ya usaba la noradrenalina pero sin atreverse a señalarla como el mediador químico, pues en ese momento no se sabía si con certeza existía en el organismo. También había descubierto la acción oxitócica de los extractos pituitarios, demostrando que estos eran diferentes de los principios que producían elevación de la presión arterial. Posteriormente y en colaboración con P. P. Laidlaw, identificó la histamina en tejidos animales, ya que hasta ese momento solo era conocida como una sustancia sintética y para 1918 había determinado algunos de sus efectos fisiológicos, como la vasodilatación y participación en el shock anafiláctico.
Jugó el papel clave al diseñar las normas internacionales para las sustancias activas biológicas como las hormonas, antitoxinas y vacunas. Se le atribuye la autoría de la llamada Ley o Principio de Dale, según el cual las neuronas pueden diferenciarse en razón del neurotransmisor que liberan y el cual estuvo vigente hasta que recientemente, se describió la posibilidad de producción de otras sustancias como neuropéptidos y aminoácidos (principio de la coexistencia).
En 1913 realizó experimentos con acetilcolina, observando que su inyección intravenosa provocaba la inhibición de los latidos cardíacos, simulando los mismos efectos de la estimulación parasimpática. Sus experimentos con preparaciones musculares de sanguijuelas le aportaron información adicional sobre la liberación de acetilcolina en la unión neuromuscular. Pero durante esta época ese descubrimiento no tuvo mayor trascendencia ya que tampoco se sabía que la acetilcolina existiera en el organismo. Ciertamente, sus trabajos podían considerarse como los albores de la hipótesis química y no transcurrió mucho tiempo hasta que otros neurofisiólogos rápidamente rechazaran tales sugerencias: Eccles como ya vimos y John Fulton, también discípulo de Sherrington y editor de la revista Journal of Neurophysiology, quien textualmente se expresó acerca del asunto del siguiente modo: “…La idea de un mediador químico liberado en las terminales nerviosas y actuando sobre una segunda neurona o un músculo parece insatisfactoria en muchos aspectos…”.
El problema que enfrentaba Dale (Figura 6) era como probar que la sustancia química que habían identificado después de estimular un nervio y obtener una contracción, provenía del nervio mismo y no de la actividad muscular. En tales circunstancias, Dale escribió: “…Si por otra parte los músculos están paralizados por el efecto del curare sobre el impulso nervioso, la estimulación de su nervio motor produce la liberación habitual de acetilcolina, aunque el músculo permanezca completamente pasivo…”. Fue de esta manera como una vez más utilizando curare se pudo comprobar que la acetilcolina encontrada en la sangre proveniente del músculo contraído se originaba completamente en el nervio. Una vez alcanzado este punto, en 1936, demostró que la cantidad de acetilcolina liberada por un solo impulso nervioso era del orden de 10-15 gramos y que su administración a través de los vasos aferentes producía una contracción muscular.

Figura 6. Henry Dale: retrato.
Henry Dale había nacido en Londres el 9 de junio de 1875, su madre fue Frances Ann Hallet y su padre Charles James Dale fue un hombre de negocios. Se casó con su prima-hermana Ellen Harriet Hallet Dale en 1904 con quien tuvo tres niños, de los cuales su hija mayor, Allison Sarah fue la esposa de Lord Alexander R. Todd, premio Nobel de Química en 1957. Murió en Cambridge el 23 de julio de 1968.
Inicialmente fue educado en el Tollington Park College en Londres y posteriormente asistió al Leys School en Cambridge como paso previo para completar sus estudios de pregrado en ciencias naturales en el Colegio de la Trinidad. Trabajó durante varios años con Langley, Gaskell y F. G. Hopkins hasta 1900 antes de completar su graduación como médico y de recibir su título en Cambridge. A partir de este momento se dedicó a la investigación, comenzando con Ernest Starling en el University College y siguiendo con Paul Ehrlich en Alemania hasta regresar a los laboratorios de la farmacéutica Burroughs-Wellcome, que posteriormente dirigió. A pesar de los consejos en contrario para trabajar en un empleo comercial, decidió aceptar la oferta y asegurarse un lugar propio de trabajo con un ingreso estable que a la vuelta de 10 años le produjo el reconocimiento como la figura más grande de la farmacología experimental. En 1914 fue designado Director del Instituto Nacional de Investigaciones Médicas y durante la segunda guerra mundial fue consejero científico del Gabinete.
Fue precisamente en aquellos mismos laboratorios donde estableció a través de una serie de experimentos que una de las dos sustancias aisladas por Loewi en 1921 y que este llamara Vagusstoff (sustancia vagal) era precisamente la acetilcolina. Ambos, quienes personalmente fueron entrañables amigos y trabajaron juntos, compartieron el Premio Nobel de 1936, en cuya ocasión Dale se expresó textualmente: “…Déjeme decirles que el orgullo y el placer que siento por este premio se ensancha por el hecho de compartirlo con mi viejo e íntimo amigo Profesor Loewi a quien conocí como un joven hace 35 años…”. Dale que también había aislado la misma sustancia en el bazo de grandes animales, concluyó que en las especies de laboratorio sus concentraciones eran muy escasas o se destruía muy rápido. De hecho reconocía la posibilidad de que su “…extraordinaria evanescencia sugería que una esterasa probablemente contribuía con su rápida desaparición de la sangre…” (Conferencia Nobel). En esta misma ocasión añadió: “…La secuencia de los eventos incluye dos hechos: el primero es un depósito estrechamente ligado a...las terminales del nervio motor, en el cual se pueda almacenar la acetilcolina, protegerla de su destrucción y desde donde igualmente pueda ser liberada con la llegada de un nuevo impulso. El Prof. Loewi ya ha mencionado esta evidencia y nosotros hemos demostrado que toda la droga que se obtiene por extracción desaparece cuando las fibras nerviosas se degeneran. El segundo factor es un mecanismo muy rápido para su destrucción que la haga desaparecer en los pocos milisegundos que dura el período refractario y que Loewi identificara en el músculo cardíaco…suponiendo que se encuentra concentrada en inmediata relación con el lugar de su liberación…”. Nada más cercano a la realidad que hoy conocemos.
b) Consolidación
Otto Loewi siempre reconoció recordar el último párrafo de una comunicación de Elliott en la cual sugería la posibilidad de que una sustancia fuera liberada después de estimular los nervios simpáticos (J. Physiol., 1904;31:20), así como aquel otro donde Dixon demostraba algo parecido al estimular el vago (Med Mag 1907;16:454). Loewi personalmente describe de qué manera en 1921 obtuvo sus propias pruebas para explicar tales hallazgos.
Para esa época en su laboratorio se venían haciendo experimentos con corazones escindidos de conejos y ranas a fin de estudiar entre otras drogas, la digital. Loewi mismo refiere que una noche y mientras trataba de conciliar el sueño se le ocurrió una idea. Se levantó y la describió en un pedazo de papel que dejó en su cama. A la mañana siguiente solo guardaba débiles recuerdos de lo acontecido y parecía como si la brillante idea se hubiese convertido en un fantasma de su imaginación. Busco el papel y trató de descifrar lo que había escrito, describiéndolo textualmente de la siguiente manera: “…Me levanté inmediatamente y me fui al laboratorio donde realicé el sencillo experimento con el corazón de una rana, de acuerdo al diseño de la noche anterior…”. Durante el mencionado ensayo había estimulado los nervios del corazón de una rana y como consecuencia se debía haber liberado una sustancia que al ser transferida al medio de perfusión y luego a otro corazón, producía en este una reacción exactamente similar a la que se obtuviera por la estimulación de sus propios nervios. En conjunto quedaba probado que los nervios no actúan directamente sobre el corazón, sino que el químico liberado era responsable de los cambios producidos en la función cardíaca. Posteriormente llegaría a demostrar que el efecto descrito podía ser antagonizado por la administración de atropina (Pflügers Arch. Ges. Physiol, 1921; 189: 239) y era de breve duración (Pflügers Arch. Ges. Physiol, 1926; 214: 678), propiedades que solo parecían corresponderse con la acetilcolina (idem: 689).
Más tarde y en forma determinada asegura que: “…la liberación de una sustancia por el nervio es siempre la expresión de una función positiva: la activación. Esto comprueba que el efecto directo de la estimulación de todos los nervios, activando o inhibiendo, representa su verdadera función y por tal motivo es que se libera la sustancia…”.
El investigador continúa con sus pensamientos y entonces se pregunta desde dónde exactamente es liberada la acetilcolina y él mismo se responde que esa función solo puede tener lugar en las terminaciones nerviosas o en el órgano efector. Pero dada la circunstancia de que la acetilcolina no está presente en la sangre ni puede difundir desde otra región, ello le confirma que esa sustancia se forma en el nervio, donde estaba presente aún en condiciones de reposo.
Otto Loewi (Figura 7) había nacido el 3 de junio de 1873 en Frankfurt-am-Main, hijo de Jacob Loewi un mercader en vinos y de Anna Willstätter, quienes le indujeron el interés por las artes y las ciencias, en consecuencia su inclinación inicial fueron la pintura y la música. En 1908 contrajo matrimonio con Guida Goldschmiedt, hija del Dr. Guido Goldschmiedt, quien era profesor de química en Praga y después en Viena. Tuvieron cuatro hijos: Hans, Victor, Guido y Anna. Loewi. En 1946 se hizo ciudadano americano y murió el 25 de diciembre de 1961.
En 1891 terminó sus estudios sobre los clásicos mientras realizaba largos viajes con tal propósito y particularmente se interesó en la historia del arte. Pero a esta altura sus padres le sugirieron estudiar una profesión y la posibilidad de que llegara a convertirse en “doctor”. Solo en el verano de 1893 se dedicó seriamente a la medicina al aprobar su examen de admisión. Recibió el grado de doctor de la Universidad de Strassburg, siendo su tesis dirigida por el famoso Prof. Schmiedeberg, llamado el Padre de la Farmacología. Durante los años 1897 y 98 fue asistente del afamado clínico von Noorden, pero impresionado por la elevada mortalidad causada por la tuberculosis, sin que pudiera contar con un tratamiento eficaz, decidió dedicarse a la investigación en materias básicas y en ese momento se hizo asistente del Prof. Meyer en la Universidad de Marburg-an-der-Lahn. Durante sus primeros años se dedicó al campo del metabolismo, pero en 1902 pasó algún tiempo en el laboratorio de Starling y su cuñado Bayliss en Londres donde por primera vez conoció a Dale. En 1904 ya era profesor de farmacología en Viena, para que finalmente ascendiera en 1909 como Jefe de la Cátedra de Farmacología en la Universidad de Graz.
Posteriormente salió del país y pasó un breve período en la Universidad Libre de Bruselas y en el Instituto Nuffield en Oxford. Más tarde, en 1940 aceptó su designación como profesor e investigador de farmacología en el Laboratorio George Wallace de la Universidad de Nueva York. En reconocimiento a sus descubrimientos, se hizo acreedor al Premio Nobel de 1936, compartido con Dale.

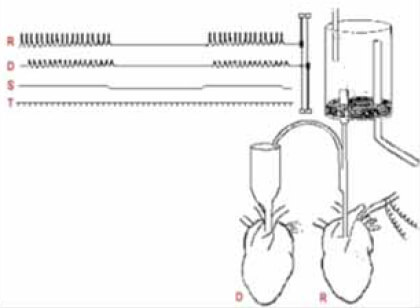
Figura 7. Otto Loewi: retrato y reproducción esquemática de su famoso experimento de perfusión.
Otra prominente figura de las ciencias y en particular de la investigación sobre los neurotransmisores fue Wilhelm Siegmund Feldberg. Como ya hemos referido someramente, para 1904 Elliot era solo un estudiante de Cambridge pero ya había descrito el parecido entre las respuestas a la inyección de adrenalina con la estimulación de los nervios simpáticos. Más adelante, también dejamos constancia de que 1909 Dixon había propuesto que los nervios parasimpáticos liberaban una sustancia parecida a la muscarina, mientras que Dale en 1914 describía, no solo a la acetilcolina como un agente parasimpaticomimético sino también la presencia de la colinesterasa en la sangre y en los años siguientes Lowei anunciaba su descubrimiento de la sustancia vagal.
Esta sucesión de hechos científicos parece haber incitado a Feldberg para investigar la liberación de acetilcolina después de la estimulación nerviosa. Una seria limitación para realizar este proyecto era la carencia de un método confiable para su determinación. Con el objeto de obviar esta dificultad utilizó inicialmente la técnica de “eserinizar” al animal y obtener las muestras del órgano investigado, siendo la lengua y posteriormente el corazón los primeros en ser utilizados. Pero uno de sus experimentos cruciales consistió en la inyección intrarterial de acetilcolina en una preparación para provocar una contracción muscular y conjuntamente recolectar muestras después de la estimulación eléctrica de sus nervios. A partir de este momento volcó su atención para descubrir la síntesis de la acetilcolina y en el curso de esta investigación pudo definir la acción de la colinacetilasa en extractos de tejido cerebral desecado.
Feldberg diseñó varias técnicas para localizar el sitio de acción de algunas drogas y con este propósito también utilizó el curare, pero esta vez, no para provocar bloqueo neuromuscular sino más bien desencadenar convulsiones y descargas en el corticograma cuando se lo inyectaba en el ventrículo lateral del gato. Se interesó en los mecanismos de producción de la fiebre, describiendo la elevación que sigue después de administrar 5-hidroxitriptamina y la disminución que se observa por acción de la adrenalina. Mostró gran interés por la histamina y sus trabajos en esta área lo llevaron al descubrimiento de la llamada sustancia de reacción lenta de la anafilaxia. Participó activamente durante la guerra mundial demostrando que la toxicidad de los gases venenosos se debía a su efecto supresor sobre algunas acciones de la colinesterasa.
Feldberg nació en Hamburgo en el seno de una familia de clase media, el 19 noviembre de 1900. Estudió medicina en Heidelberg, Munich y Berlín, graduándose en 1925. Al contraer matrimonio se trasladó con su esposa a Inglaterra, donde curso estudios con Langley en Cambridge y luego con Dale en el Instituto Nacional de Investigaciones Médicas de Hampstead. En 1927 regresó al Instituto Fisiológico de Berlín. Su primera esposa Katherine fue traductora, profesora y experta en literatura, pero murió en 1976. Posteriormente se casó con Kim O´Rourke, quien hizo un formidable trabajo para publicar sus obras, pero desafortunadamente falleció de manera trágica cuatro años más tarde. Su hijo murió en 1960 y su hija se dedica a la cardiología pediátrica en Seattle. Murió en Londres el 25 de octubre de 1993. Después de la guerra había utilizado los fondos que recibió como compensación, así como la pensión otorgada en su condición de profesor emérito, para la creación de la Fundación Feldberg dedicada al intercambio científico.

Figura 8. Wilhelm S. Feldberg: retrato y portada de una de sus obras.

A finales de la década de 1940, Bernard Katz secundado por Alan Hodgkin demostraron el papel del sodio en la generación del potencial de acción y desarrollaron la técnica del “clamp voltage” para el estudio de las corriente iónicas. En una segunda ocasión, pero en compañía de P. Fatt reveló el mecanismo por el cual el transmisor químico liberado en las terminales nerviosas producía una corriente eléctrica en en intensidad y tiempo al potencial de acción en la placa motora. Estos resultados lo llevaron a concluir que la acetilcolina alteraba la configuración de algunas moléculas en la membrana celular permitiendo el flujo iónico y la generación de corrientes eléctricas.
En un principio los investigadores de ese momento seguían aceptando la transmisión eléctrica a nivel del sistema nervioso central, de manera que los descubrimientos de Katz circunscritos exclusivamente para la unión neuromuscular, eran comparativamente menores. Aún no se conocía ningún neurotransmisor en otra localización y la existencia de la membrana celular era solo una proposición, nadie la había visto. Por añadidura, los receptores y canales eran únicamente conceptos que apenas representaban la mejor explicación experimental, pero tampoco habían sido vistos. Toda esta situación puede resumirse en sus propias palabras tomadas de la conferencia inaugural en el University College en 1952: “…Yo no me atrevería a traspasar el dominio de la mente, acerca de la cual tanto se ha dicho y tan poco se puede decir y habiéndoles dejado saber mi disposición pedestre, yo espero que ustedes me dejarán detener en este punto…”.
Katz soñaba con la posibilidad de poner en evidencia los fenómenos producidos por una sola molécula de transmisor en la medida que esta se uniera al receptor y daba origen al flujo de iones. Propuso que la super imposición de una despolarización estable debía incluir esas pequeñas fluctuaciones de voltaje y cuyo mecanismo íntimo debía ser la apertura y cierre de los canales activados por la acetilcolina. Con la ayuda de una herramienta matemática y estadística apropiada que se llama “análisis de la fluctuación” se pudo entonces estimar cuantitativamente la intensidad y duración del flujo de la corriente a través de un solo canal e igualmente fue capaz de seguirla en tiempo real a medida que las “compuertas” se abren y cierran.
Pasado algún tiempo, el mismo Katz (Figura 9) describió parte de sus investigaciones de la siguiente manera: “…Nosotros estábamos seguros de que los potenciales en miniatura eran un fenómeno normal y que no era producido por lesión alguna de las terminaciones nerviosas motoras. Con el tiempo llegaron a convertirse en un descubrimiento de extraordinario interés que nos trajo muchos años de ocupación seria y entretenimiento para mí y mis colegas…Debo reconocer que la exploración experimental de la naturaleza de estos minipotenciales me proporcionó mucho disfrute…” (B.Katz: Planning and following the unexpected in scientific research. Creativity. Res J. 1994;7:225-238).
Bernard Katz nació en Leipzig el 26 de marzo de 1911, de padres rusos y polacos. Siendo su padre ciudadano del Imperio zarista, se vio desprovisto de patria después de la revolución y esto lo indujo a emigrar para Alemania. Katz fue educado en el König-Albert-Gymnasium y continuó estudiando medicina en la universidad de su ciudad natal. Se graduó en 1934 y voló a Gran Bretaña en febrero de 1935. Entró a trabajar en el University College de Londres, bajo la tutela de A. V. Hill. Terminó su doctorado en 1938 y ganó una beca para estudiar al lado de J.C. Eccles en Australia. Se naturalizó en 1941, uniéndose a las Fuerzas Aéreas Australianas en 1942 para servir como oficial de Radar en Nueva Guinea. En 1970 recibió el Premio Nobel que compartió con Julius Axelrod y Ulf von Euler y en 1978 fue declarado profesor emérito, falleciendo en Londres el 20 de abril de 2003.
Fue en Australia donde conoció a quien fue su esposa, Marguerite Penly (Rita) y juntos regresaron a Londres después de la guerra. Le gustaba la música, particularmente Verdi y jugaba al ajedrez. Rita desarrolló una prolongada enfermedad y falleció en 1999. A los Katz les sobrevivieron sus hijos David, médico y Jonathan músico y profesor, así como tres nietos: Joseph, Stephanie y Judith. En su momento culminante, Katz quien en el curso de sus investigaciones registraba los eventos de la placa motora con la utilización de electrodos intracelulares, también demostró la presencia de pequeñas diferencias de potencial, de 1 mV aproximadamente que se presentaban una vez por segundo y podían ser abolidos por la acción del curare, cuya naturaleza era desconocida y que consideró como fenómenos espontáneos. Fue así como se estableció que estos potenciales en “miniatura” tenían su origen en la liberación espontánea del neurotransmisor desde las terminaciones nerviosas y una vez más dejó claro el papel del curare en la explicación de la transmisión neuromuscular.
BIBLIOGRAFÍA
- Reveu des cours scientifiques de la France et de l’etranger. Deuxiéme Anne. Paris. Germere Bailliére, Libraire et Editeur. 1864 – 1865.
- Cousin MT. Vulpian and not Claude Bernard proponed the hypothesis of the motor end-plate as the site of action of curare. Anesthesiology. 2002;97:527-528.
- Mahele A-H. Receptive substances: John Newport Langley (1852-1925) and his path to receptor theory of drug action. Medical History. 2004;48:153-174.
- Anglade P, Tsuji S. Hundredth anniversary of the synapse. I: a short history of the milestones in synapse research. Zoological Science. 2004;14:533-537.
- Cuthbert AW. The man who never was: Walter Ernest Dixon FRS. Br J Pharmacol. 2001;133:945-950.
- Sherrington CS. The Integrated Nervous System (Cap 14) En: Finger S, editor. Minds behind the Brains. A History of the Pioneers and Their Discoveries. Londres: Oxford University Press; 2000.p.217-237.
- Loewi O, Dale H. The Discoverers of Neurotransmitters (Cap 16) En: Finger S, editor. Minds Behind the Brains. A History of the Pioneers and Their Discoveries. Londres: Oxford University Press; 2000.p.259-279.
- Axelrod J. Journey of a late blooming neuroscientist. J Biol Chem. 2003;278:1-13.
- Sabatini RME. Neurons and synapses: The history of its discovery. Brain & Mind Magazine. 2003;17:April – July.
- Bennett MR. History of the synapse. Nueva York: Harwood Academic Publishers; 2001.
- Fletcher WM. John Newport Langley: In memoriam. J Physiol. 1926;61:1-27.
- Bisset GW. Wilhelm Feldberg (1900-1993). Br J Pharmacol. 1994;111:649-651.
- Todman D. John Eccles (1903-97) and the experiment that proved chemical synaptic transmission in the central nervous system. J Clin Neurosc. 2008;15: 972-977.
- Eccles JC. An electrical hypothesis of synaptic and neuromuscular transmission. Ann N Y Acad Sci. 1946;47:375-602.
- Todman D. Henry Dale and the discovery of chemical synaptic transmission. Eur Neurol. 2008;60:162-164.
- Tansey EM. Henry Dale and the discovery of acetylcholine. Compt Rend Biolog. 2006;329:419-425.
- Feldman S. Poison arrows. Londres: Metro Publishing Ltd.; 2005.
