Historia de la enfermedad de Chagas en el mundo y en Venezuela (Segunda parte)
Dr. José Antonio O'Daly
Individuo de número de la Academia Nacional de Medicina, Caracas, Venezuela.
(Trabajo presentado en el XI Congreso de la Sociedad Venezolana de Historia de la Medicina realizado en Caracas, del 22 al 24 de octubre de 2019)
Situación actual
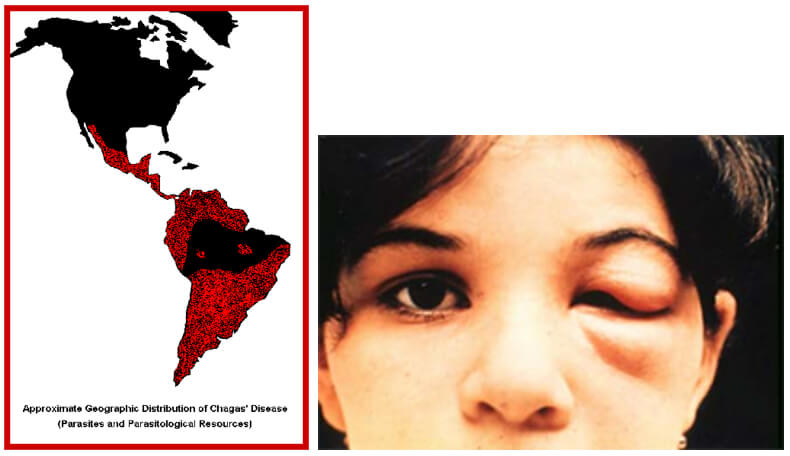
Actualmente se estima que en Latinoamérica existen de 7-8 millones de personas infectadas con T. cruzi y que más de 25 millones están en riesgo de contraer la ECh [1, 73], solamente en el año 2008 se reportaron más de 10.000 muertes por ECh [73]. Desde la década de 1990 las iniciativas multinacionales han conducido a una reducción significativa en el número de casos agudos de ECh, así como también en la presencia de Triatomineos domésticos en muchas regiones endémicas en Latinoamérica [1]. En Centroamérica el rociamiento con insecticidas residuales ha eliminado el R. prolixus [74]. A pesar de estos logros en el control del parásito y del vector han aparecido nuevos desafíos, incluyendo nuevos brotes de ECh en la cuenca amazónica, una región que se creyó previamente libre de la enfermedad, debido a transmisión oral por alimentos contaminados [74, 75, 76], y la transmisión activa de ECh en la región del Chaco Boliviano a pesar de los programas de control de vectores en progreso desde el año 2000 [77]. Otro problema es la aparición de vectores Triatomineos resistentes a los insecticidas en el Gran Chaco una región localizada al oeste del rio Paraguay y al este de los Andes [78]. A pesar de los numerosos esfuerzos en las décadas pasadas no se han desarrollado drogas para el tratamiento de la ECh crónica, y de acuerdo a las nuevas recomendaciones hechas en 2005 y 2007 el tratamiento con Nifurtimox y Benznidazole está indicado para pacientes con infección aguda así como también para pacientes menores de 18 años de edad con infección crónica, así como también en pacientes inmunosuprimidos [79, 80]. Los costos para el tratamiento y prevención de la ECh son otro desafío y son una carga muy alta para los sistemas de salud en los países afectados. En Colombia solamente los costos para la atención médica de los pacientes con ECh son de US $ 267 millones y para el rociamiento con insecticidas para el control, de vectores de US $ 5 millones, [81]. (Fig 14). La ECh se está transformando en un problema global debido a las migraciones de personas infectadas con T. cruzi desde los países endémicos a Norteamérica, Europa y la región del Pacifico Oeste. El número total estimado de pacientes con ECh fuera de Latinoamérica es más de 400.000, siendo USA el país más afectado con 3-4 partes de todos los casos [82, 83]. En Europa solamente el número de pacientes con miocardiopatía crónica chagasica se estima en 54.000 casos [83].
Biología Molecular T. cruzi
Encontramos que varias proteínas del suero fetal de ternera (SFT) estimulan la división y la incorporación de 3HTimidina en T. cruzi mediante su interacción con la membrana celular de los tripanosomas. En nuestro primer medio de cultivo [84] , [85] el T. cruzi presentó un ciclo con 100% de epimastigotes en la fase exponencial de crecimiento al día 12 y 88% de tripomastigotes en la fase estacionaria al día 16 de cultivo. Cinco proteínas del SFT estimuladoras de la división del T. cruzi mostraron los siguientes pesos moleculares: Proteína 1: 100.000 Da; proteína 2: 76.000 Da; proteínas 3, 4 y 5: 68.000 Da [86]. Las proteínas cambiaron su estructura durante la fase logarítmica de crecimiento del T. cruzi, base de la autoinmunidad en la ECh [87]. El complejo mitocondria-kinetoplasto en los epimastigotes mostro una nueva estructura formada por un patrón de canales con una estructura como un cristal en el citoplasma y también otra nueva estructura al final del flagelo para fijar el T cruzi a la superficie de los frascos de cultivo plásticos, y a las membranas celulares de las células de los mamíferos [88]. La inmunoglobulina IgG de cabra con actividad específica anti-proteínas del SFT que estimulan la división de T cruzi marcada con fluorocromos se localizó en el citoplasma de T cruzi. Usando las proteínas que estimulan la división del parasito en el SFT marcadas con 125I en el medio de cultivo, estas también se encontraron en gránulos citoplasmicos en el T cruzi mediante autoradiografía. Igualmente las proteínas del SFT que estimulan la división marcadas con oro coloidal, se encontraron en las membranas, citostoma, libres en el citoplasma y en gránulos citoplasmicos, probablemente lisosomas algunos abiertos al núcleo del parasito en división en contacto con la membrana perinuclear enviando péptidos que inducen la división del DNA y la división celular en los tripanosomas [89]. La fracción insoluble de las membranas de T. cruzi, L. donovani y L. mexicana inducen la lisis de glóbulos rojos de mamíferos y células Vero. El agente lítico es resistente a la pronasa, a la tripsina y al aumento de la temperatura, su acción tiene un pH óptimo de 6.5 y está presente en la fase lipídica de los extractos de T. cruzi; su acción lítica es inhibida por la albumina [90] , [91]. El T. cruzi tiene un patrón característico de aminoácidos libres diferente al de las células de mamíferos, producidos por su actividad proteolítica, formado en un 75% por acido glutámico, prolina, glicina y alanina, lo cual debe tenerse presente en los procesos de purificación de antígenos en los extractos de parásitos para la producción de vacunas. La actividad proteolítica del parasito tiene un óptimo entre pH 4.5 a 8.0 y es inhibida por inhibidores de enzimas proteolíticas tales como ZPCK, TPCK, PMSF, and TLCK [92]. Estos resultados demuestran que la inyección de parásitos o sus fracciones, debe considerar la destrucción de las proteínas antigénicas en un periodo de tiempo muy corto, inhibiendo la respuesta inmune protectora contra los parásitos virulentos. Con los valores obtenidos de la concentración de aminoácidos libres en T cruzi producto de su actividad proteolítica a las 24 horas en los extractos totales del parasito preparamos un nuevo medio de cultivo enriquecido (medio O’Daly) que permitió el crecimiento de epimastigotes de T cruzi entre 26° - 37° C. donde analizamos los requerimientos nutritivos de T cruzi a diferentes temperaturas con SFT o en péptidos purificados del SFT. Un péptido compuesto por un ácido glutámico, dos alaninas y una lisina substituyo al SFT en el crecimiento de los tripanosomas en el medio O’Daly [93, 94]. Ratones homocigotos C3H/He y C57Bl/6 infectados con 500 tripanosomas de la cepa Y de T. cruzi mostraron parasitemia, aumento del bazo y de los ganglios linfáticos, parásitos intracelulares en el corazón y aumento en la concentración de la IgG y la IgM en el suero. En el punto mayor de la parasitemia los 17 días post-infección, se encontró una disminución marcada de la respuesta de los linfocitos a mitogenos de linfocitos T y B a los antígenos del parasito y a las células formadoras de placas a glóbulos rojos de carnero [95]. Tripomastigotes sanguíneos de la cepa Y de T. cruzi inhibieron la respuesta de mitógenos T y B en células nucleadas de bazo de ratones C3H/He, C57Bl/6 y BALB/cJ, mientras que epimastigotes de cultivo de la misma cepa de T cruzi cultivados en medio O’Daly que permite el crecimiento de los parásitos a 26°, 30°, 34° y 37°C indujeron un efecto estimulador muy fuerte, aun mayor que el efecto de los mitogenos solos. Tanto el efecto estimulador como el efecto inhibidor mostraron una relación dosis-respuesta. El efecto estimulador dependió de la temperatura, los índices de estimulación fueron mayores al aumentar la temperatura de los cultivos [96]. Los sobrenadantes de células de bazo de ratones C3H/He y C57Bl/6 infectados con T. cruzi mostraron proteínas hidroinsolubles de peso molecular entre 14.000-15.000 daltons, pI 6.6, capaces de inducir un efecto inmunosupresor de la respuesta blastogenica a mitogenos in vitro. El efecto inhibidor atravesó de la barrera haplotipica H2K-H2b, su actividad fue dependiente de la dosis, su producción se relacionó con el periodo de infección mostrando un máximo a los 17 días post-infección por los esplenocitos infectados. Es un producto del huésped infectado y actúa a través de su interacción con macrófagos normales transformándolos en inmunosupresores. Una proteína inmunosupresora con igual pI de 6.6, ausente del suero normal se detectó en el suero de los animales infectados [97]. A fin de detectar antígenos de tripanosomas en la superficie de linfocitos de ratones infectados se usaron antisueros anti-epimastigotes y anti-tripomastigotes de T. cruzi. Solo el suero anti-tripomastigotes reconoció antígenos expresados transitoriamente en las membranas de esplenocitos de animales infectados. El número o la conformación estructural de los receptores de Concanavalin A fueron también afectados y se vio una correlación clara entre estos dos tipos de cambios en la membrana y la inmunosupresión a mitogenos y a glóbulos rojos de carnero (GRC) encontrada en los ratones infectados. Los animales re-infectados no mostraron evidencias de proliferación de tripanosomas en la sangre o en los tejidos, ni tampoco antígenos de tripomastigotes en los esplenocitos, pero presentaron una inmunosupresión transitoria menos intensa, medida por la respuesta a mitogenos y GRC lo cual sugiere que el sistema inmune activado una primera vez puede eliminar el nuevo inoculo parasitario antes de que el huésped este inmunosuprimido y también que la liberación de antígenos fuertes inmunosupresores por los tripomastigotes induce el nuevo estado menos intenso y transitorio de supresión [98]. Epimastigotes de T. cruzi cambian el patrón de proteínas de superficie cuando la temperatura de crecimiento en el cultivo aumenta de 30° a 34° C. El reto de los ratones con tripomastigotes sanguíneos produjo parasitemias altas cuando los animales fueron inmunizados con proteínas de superficie de epimastigotes cultivados a 30° C y con sedimentos de epimastigotes extraídos con Nonidet P-40 cultivados a 34° C. Por el contrario se observaron parasitemias bajas después de la inmunización con proteínas de superficie de epimastigotes cultivados a 34° C o de sedimentos de epimastigotes extraídos con Nonidet P-40 y cultivados a 30° C. La parasitemia menor con el mayor tiempo de sobrevida y ausencia de inmunosupresión se observó después de la inmunización de los ratones con el producto extraído con el detergente no-iónico NP-40, de epimastigotes cultivados a 30° C y tratados con Tosyl-L-lysine-chloromethyl ketone [99].
Productos secretados por L. (L) amazonensis (La); L. (L) venezuelensis H16 y H17 (Lv16, Lv17); L. (V) brasiliensis H27 y H2 (Lb27,Lb2); L. (L) chagasi (Lch); L. (L) pifanoi (Lp) y L. garnhami (Lg) fueron caracterizados en un medio de cultivo químicamente definido [93] (Medio O’Daly) donde se encontraron bandas de proteínas y glicoproteínas reveladas en geles coloreados con los siguientes números, Coomassie (13.8±1.5), Plata (10.2±3.3), PAS-Schiff (7.4±2.5) y ConA(5.33±1.9). La inmunización con productos secretados de Lg, Lv16, Lp y con amastigotes de cultivo tratados con TLCK y extraídos con NP-40 indujeron protección completa contra la infección con amastigotes virulentos desde la semana 14 a la semana 24 de seguimiento. ANOVA unidireccional entre Lg, Lv16, Lp, vacuna polivalente (VP) y control no inmunizado (CNI) mostraron p<0.001. Lch, Lv17, Lb and La, también mostraron regresión de las lesiones con solo un 5%-10% de aumento en la almohadilla plantar comparadas con el CNI. La reacción de hipersensibilidad retardada (DTH) fue mayor con los productos secretados de Lg, Lv16, Lp y la VP, lo cual se correlaciona con la ausencia de lesiones después de la infeccion. Lch, La, y Lb también mostraron valores de DTH positivos pero menores que la VP. Los immunoblottings con sueros de ratones inmunizados con Lg, Lv16, y Lp mostraron bandas densas de 83 kDa y de 50 a 55 kDa. Después de la infeccion de los ratones inmunizados ocurre un suiche en las células B, los anticuerpos IgG ahora reconocen productos secretados de bajo peso molecular entre 27-33 kDa y 23-26 kDa coincidentes con la regresión completa de las lesiones lo cual no ocurrió en ratones infectados con Lch, Lv17, Lv16, and Lp donde las lesiones persistieron [100].
Proteínas de superficie, secretadas y totales de T. cruzi y T. rangeli crecidos en medios sintéticos, químicamente definidos (medio O’Daly) [93] se usaron como antígenos para el diagnóstico de la ECh por ELISA e Immunoblottings. El suero de pacientes con miocardiopatías Chagasicas y no Chagasicas, de donantes de sangre, de controles sanos, y de pacientes con otras enfermedades tropicales mostraron extensas reacciones cruzadas entre los antígenos de T. cruzi y T. rangeli, los cuales mostraron patrones muy similares en los immunoblottings. El patrón antigénico con sueros chagasicos es muy específico y diferente a los patrones con otras enfermedades tropicales. Las proteínas secretadas por T. cruzi así como también las infecciones por T. rangeli juegan un papel importante en la patogenia de la ECh [101].
Patogénesis de la Enfermedad de Chagas
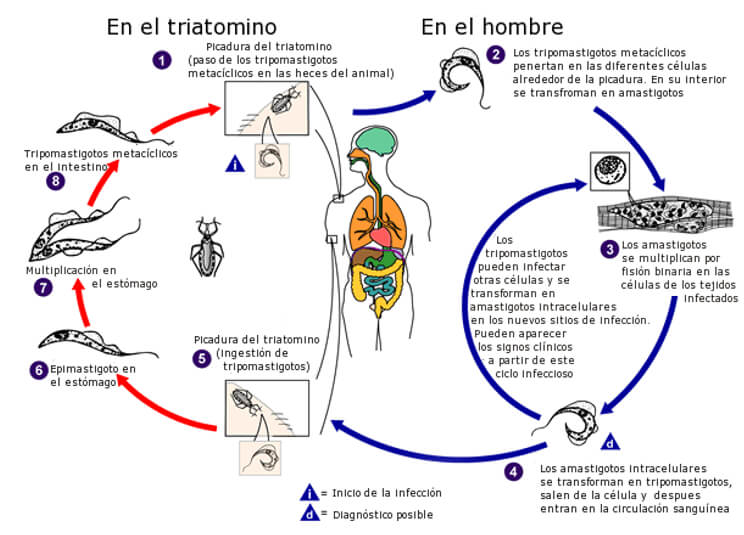
El ciclo de vida del T. cruzi nos ilustra sobre la versatilidad del parasito en colonizar vertebrados e invertebrados (Fig 13) en un área geográfica extensa en Norte, Centro y Sur América (Fig 14). La patogénesis de la ECh parece estar relacionada a una mutación inducida por el parasito en el genoma del vertebrado infectado. Se ha detectado transferencia horizontal de los minicírculos del DNA del kinetoplasto (kDNA) al genoma del huésped mamífero [102] , [103] , [104] , [105] , [106]. Más aun, una secuencia truncada de kDNA se encontró en el genoma de un baboon (Papyo hamadryas) en los flancos del DNA del huésped [107]. Transferencias (Blots) con el método de Souther del DNA genómico de corazón, musculo esquelético e intestino de un conejo con ECh crónica hibridizaron con una sonda de kDNA de T.cruzi. Se ha sugerido que la infeccion persistente por T cruzi puede ser una fuente acumulativa de mutaciones en el kDNA del parasito [107]. En la actualidad los tratamientos no son satisfactorios. Existen 40 especies de Triatomineos que pueden ser portadores de T. cruzi y transmisores potenciales de la infeccion [19], las cuales se adaptaron al área peri-domiciliar en los ecosistemas secos de América Central y América del Sur y son los principales transmisores de la infeccion (Fig 13) responsables de 10-12 millones de casos humanos de ECh [4, 108]. Luego siguen en importancia epidemiológica las especies de Rhodnius que habitan en climas húmedos tropicales y las especies de Panstrongylus que son ubicuas [4, 108]. Después de una comida con sangre los Triatomineos se hinchan y los tripomastigotes metacíclicos infecciosos pasan a las heces del vector. Los tripomastigotes entran al cuerpo cuando el individuo se rasca la piel en la herida producida por la parte bucal del insecto o también a través de una mucosa permisiva o la conjuntiva ocular (Fig. 15). Al entrar en el cuerpo (Fig. 13), los tripomastigotes invaden a los histiocitos, sobreviven a las vacuolas parasitoforas y evaden el ambiente hostil para permanecer libremente en el citoplasma celular de la célula huésped. En la puerta de entrada, tanto en la piel como en la conjuntiva ocular, clínicamente se observa un proceso indurado inflamatorio crónico, típico de una reacción de sensibilidad retardada (DTH). Los flagelados intracelulares se redondean y se transforman en amastigotes que sufren muchos ciclos de multiplicación por fisión binaria, los cuales posteriormente se diferenciarán en tripomastigotes móviles (Fig. 13) que al romperse la célula llegaran al torrente sanguíneo para infectar otras células en el cuerpo del mamífero. Las primeras investigaciones en la ECh mostraron una relación directa entre la densidad de los receptores en la membrana del huésped infectado y la carga parasitaria en los tejidos [109]. Cada célula en el cuerpo humano excepto las neuronas puede ser colonizada por el T. cruziin vivo. La infeccion por T. cruzi persiste en el cuerpo humano toda la vida, comienza por la invasión tisular de las células reticulo-endoteliales locales, donde posteriormente el parasito se multiplica, penetra el endotelio vascular de los pequeños vasos intraparenquimatosos como los capilares y precapilares hasta alcanzar la circulación sistémica e invadir varios tejidos de acuerdo a su tropismo tisular, fundamentalmente musculo esquelético, y cardiaco, sistema digestivo, órganos linfohematopoyeticos, y sistema nervioso (Fig. 16 y 17). La disfunción endotelial dispara la cascada de eventos subcelulares que conducen a la inflamación del miocardio, pericardio y endocardio con la aparición progresiva de cicatrices en la matriz extracelular. La acción más evidente del T. cruzi es el bloqueo de las células presentadoras de antígenos a través de la proteína Tcsc y su inmunoglobulina, agonistas de los receptores β-adrenérgicos. La inoculación de T. cruzi en huevos embrionados de gallinas, genera pollos libres de parásitos que retienen secuencias de minicírculos de kDNA en el cuerpo, en ausencia del DNA nuclear del T. cruzi. El kDNA se integra en el genoma de las células somáticas y germinales, desde donde es transmitido verticalmente a la progenie que al ser cruzados entre sí, progenie positiva para kDNA. Las mutaciones en el kDNA se detectan principalmente en regiones codificadas o codones, en varios cromosomas. Es de hacer notar que los pollos con mutaciones en kDNA, desarrollan cardiomegalia con una miocarditis inflamatoria similar a la que se ve en la ECh en humanos, donde miofibrillas libres de parásitos son destruidas por las células efectoras del sistema inmunológico originando insuficiencia cardiaca. La progresión de un estado fisiológico a una condición fisiopatológica está asociada con una mutación multigenica del kDNA, y es dependiente de una proliferación clonal de células T en el modelo de los pollos, mostrando ruptura de múltiples genes, citotoxicidad acelerada anti-antígenos del animal y enfermedad de Chagas cardiaca autoinmune [104, 107].
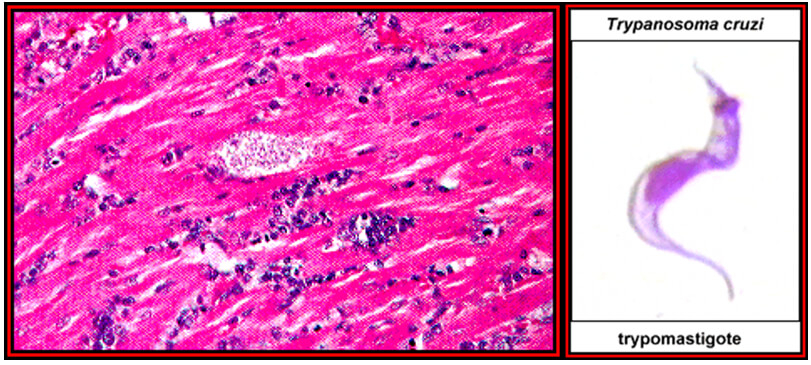
Fig 13. Ciclo del T. cruzi en el hombre (flechas azules) y en los Triatomineos, como la vinchuca y el chito (flechas rojas). Fuente: adaptado del CDC.Fig. 14 (izquierda). Población humana en riesgo de contraer la ECh. Fig. 15 (derecha). Signo de Romaña: Inflamación en el párpado izquierdo, es un indicador de la ECh aguda, debido a que se colocaron en el ojo las heces depositadas por el insecto infectado por T. cruzi en la piel. Foto de WHO/TDR.Fig. 16 (izquierda). Nidos de amastigotes en miocarditis chagasica. Fig. 17 (derecha) Tripomastigote en sangre periférica.
Conclusión
La historia de la ECh nos enseña que los cambios ambientales de origen humano son la causa primaria para la transmisión de la infección siendo la deforestación el factor más importante, al traer las poblaciones a un contacto más directo con los vectores transmisores de la enfermedad. Esto se basa en el hecho de que la transmisión de la ECh se indujo desde tiempos antiguos cuando los humanos comenzaron a limpiar la tierra para habitarla y practicar la agricultura. La minería, la industria de la madera y la urbanización unida a la deforestación, condujeron a la diseminación de la ECh en tiempos recientes. También hemos aprendido que los vectores Triatomineos tienen una habilidad increíble para adaptarse rápidamente a nuevos ambientes y a nuevos huéspedes creando ciclos de transmisión domestica entre animales y humanos cruciales para la distribución de la ECh en Latinoamérica. A través de las migraciones la ECh se ha transformado en un problema de salud global.
Referencias
D. Steverding. The history of Chagas disease. Parasit Vectors. 7 (2014), 317
C. Chagas. New human trypanosomiasis. Morphology and life cycle of Schyzotrypanum cruzi, the cause of a new human disease. Mem Inst Oswaldo Cruz. (1909), 1159–1218.
C. Chagas. A new human disease. Summary of etiological and clinical studies. Mem Inst Oswaldo Cruz. (1911), pp. 3219–3275.
A. Rassi Jr, A. Rassi, J. A. Marin-Neto. Chagas disease. Lancet. 375 (2010), 1388-1402.
A. Rassi Jr, A. Rassi, J. Marcondes de Rezende. American trypanosomiasis (Chagas disease). Infect Dis Clin North Am. 26 (2012), pp. 275–291.
A. R. L. Teixeira, N. Nitz, M. C. Guimaro, C. Gomes, C. A. Santos Buch. Chagas disease. Postgrad Med J. 82 (2006), pp. 788–798.
S. Garcia, C. Ramos, J. Senra, F. Vilas, M. Rodrigues, A. Campos de Carvalho, y col. Treatment with Benznidazole during the Chronic Phase of Experimental Chagas‘disease Decreases Cardiac Alterations. Antimicrob Agents Chemother. 49 (2005), pp. 1521–1528.
I. Maudlin, P. H. Holmes, M. A. Miles. The Trypanosomiases. Wallingford: CAB International. (2004), pp. 184.
C. A. Mendoza, E. Córdova, J. Ancca, J. Saldaña, A. Torres, R. Velásquez y col. Prevalencia de la enfermedad de Chagas en puérperas y transmisión congénita en una zona endémica del Perú. Rev Panam Salud Pública. 17 (2005), pp. 147–153.
WHO Expert Committee. Control of Chagas disease. WHO Tech Rep Ser. 905i-vi, (2002), 1-109.
C. Bern, S. Montgomery, B. Herwaldt, A. Rassi Jr, J. Marin-Neto , R. Dantas , y col. Evaluation and treatment of Chagas disease in the United States: a systematic review. JAMA. 298 (2007), pp. 2171–2181
A. Rassi, J. C. Dias, J. A. Marin-Neto, A. Rassi. Challenges and opportunities for primary, secondary, and tertiary prevention of Chagas' disease. Heart. 95 (2009), 524–534.
GBD 2015 Disease and Injury Incidence and Prevalence, Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. 388 (10053): (2016), pp. 1545–1602.
GBD 2015 Mortality and Causes of Death, Collaborators. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. 388 (10053) (2016), pp. 1459–1544.
J. L. Capinera. Encyclopedia of entomology. Dordrecht: Springer. (2008), pp. 824.
K. M. Bonney. Chagas disease in the 21st Century: a public health success or an emerging threat? Parasite. 21 (2014), pp. 11.
B. Liese, M. Rosenberg, A. Schratz. Programmes, partnerships, and governance for elimination and control of neglected tropical diseases. Lancet. 375 (2010), 67-76.
A. R. L. Teixeira, N. Nitz, M. C. Guimaro, C. Gomes, C. A. Santos Buch. Chagas disease. Postgrad Med J. 82 (2006), pp. 788–798.
R. U. Carcavallo, I. Galindez-Giron, J. Jurberg, H. Lent. Atlas of Chagas' disease vectors in the Americas. Rio de Janeiro: Editora Fiocruz, Vol 1 (1997).
E. A. Zeibig. The hemoflagellates. Clinical Parasitology: A practical Approach. Elsevier. (2013), pp. 120-123.
J. R. Stevens, H. A. Noyes, G. A. Dover, W. C. Gibson. The ancient and divergent origins of the human pathogenic trypanosomes Trypanosoma brucei and T cruzi. Parasitol. 118 (1999), pp. 107–116.
P. B. Hamilton, M. M. G. Teixeira, J. R. Stevens. The evolution of Trypanosoma cruzi: the ‘bat seeding’ hypothesis. Trends Parasitol. 28 (2012), pp. 136–141.
M. Cavazzana Jr, A. Marcili, L. Lima, F. da Silva, Â. Junqueira, H.Veludo et al. Phylogeographical, ecological and biological patterns shown by nuclear (ssrRNA and gGAPDH) and mitochondrial (Cyt b) genes of trypanosomes of the subgenus Schizotrypanum parasitic in Brazilian bats. Int J Parasitol. 40 (2010), pp. 345–355.
P. B. Hamilton, J. R. Stevens, M. W. Gaunt, J. Gidley, W. C. Gibson. Trypanosomes are monophyletic: evidence from genes for glyceraldehyde phosphate dehydrogenase and small subunit ribosomal RNA. Int J Parasitol. 34 (2004), pp. 1393–1404.
P. B. Hamilton, W. C. Gibson, J. R. Stevens. Patterns of co-evolution between trypanosomes and their hosts deduced from ribosomal RNA and protein-coding gene phylogenies. Mol Phylogenet Evol. 44 (2007), pp. 15–25.
C. A. Machado, F. J. Ayala. Nucleotide sequences provide evidence of genetic exchange among distantly related lineages of Trypanosoma cruzi. Proc Natl Acad Sci U S A. 98 (2001), pp. 7396–7401.
M. D. Lewis, M. S. Llewellyn, M. Yeo, N. Acosta, M. W. Gaunt, M. A. Miles. Recent, independent and anthropogenic origins of Trypanosoma cruzi hybrids. PLoS Negl Trop Dis. 5 (2011), pp. e1363.
J. R. Baker, M. A. Miles, D. G. Godfrey, T. V. Barrett. Biochemical characterization of some species of Trypanosoma (Schizotrypanosoma) from bats (Microchiroptera). Am J Trop Med Hyg. 27 (1978), pp. 483–491.
L. Lima, F. Maia da Silva, L. Neves, M. Attias, C. S. Takata, M. Campaner, y col. Evolutionary insights from bat trypanosomes: morphological, developmental and phylogenetic evidence of a new species, Trypanosoma (Schizotrypanum) erneyi sp. nov., in African bats closely related to Trypanosoma (Schizotrypanum) cruzi and allied species. Protist. 63 (2012), pp. 856–872.
L. Lima, O. Espinosa-Á., P. B. Hamilton, L. Neves, C. Takata, M. Campaner y col. Trypanosoma livingstonei: a new species from African bats supports the bat seeding hypothesis for the Trypanosoma cruzi clade. Parasit Vectors. 6 (2013), pp. 221.
P. B. Hamilton, C. Cruickshank, J. R. Stevens, M. M. G. Teixeira, F Mathews. Parasites reveal movement of bats between the new and old worlds. Mol Phylogenet Evol. 63 (2012), pp. 521–526.
C. V. Lisboa, A. P. Pinho, H. M. Herrera, M. Gerhardt, E. Cupolillo, A. M. Jansen. Trypanosoma cruzi (Kinetoplastida, Trypanosomatidae) genotypes in neotropical bats in Brazil. Vet Parasitol. 156 (2008), pp. 314–318.
J. Ramírez, G. Tapia-C., G. Muñoz-C., C. Poveda, L. Rendón, E. Hincapié y col. Trypanosome species in neo-tropical bats: biological, evolutionary and epidemiological implications. Infect Genet Evol. 22 (2014), pp. 250–256.
A. Marcili, L. Lima, M. Cavazzana, A. Junqueira, H. Veludo, F Maia da Silva y col. A new genotype of Trypanosoma cruzi associated with bats evidenced by phylogenetic analyses using SSU rDNA, cytochrome b and Histone H2B genes and genotyping based on ITS1 rDNA. Parasitology. 136 (2009): 641–655.
B. Stadelmann, L. K. Lin, T. H. Kunz, M. Ruedi. Molecular phylogeny of new world Myotis (Chiroptera, Vespertilionidae) inferred from mitochondrial and nuclear DNA genes. Mol Phylogenet Evol. 43 (2007): 32–48.
C. A. Flores-López, C. A. Machado. Analyses of 32 loci clarify phylogenetic relationships among Trypanosoma cruzi lineages and support a single hybridization prior to human contact. PLoS Trop Negl Dis. (2011), pp. 5:e1272.
A. C. Aufderheide, W. Salo, M. Madden, J. Streitz, J. Buikstra, F. Guhl, y col. A 9,000-year record of Chagas’ disease. Proc Natl Acad Sci U S A. 101 (2004), 2034–39.
A. Araújo, A. M. Jansen, K. Reinhard, L. F. Ferreira. Paleoparasitology of Chagas disease, a review. Mem Inst Oswaldo Cruz. 104 (Suppl. I) (2009), pp. 9–16.
A. Fernandes, A. Iñiguez, V. Lima, S. Mendonça-deS., L. Ferreira, A. Vicente y col. Pre-Columbian Chagas disease in Brazil: Trypanosoma cruzi I in the archaeological remains of a human in Peruaçu Valley, Minas Gerais, Brazil. Mem Inst Oswaldo Cruz. 103 (2008), pp. 514–516.
V. Lima, A. Iniguez, K. Otsuki, L. Ferreira, A. Araújo, A. Vicente y col. Chagas disease in ancient hunter-gatherer population, Brazil. Emerg Infect Dis. 14 (2008), pp. 1001–1002.
F. Rothhammer, M. J. Allison, L. Núñez, V. Standen, B. Arriaza. Chagas’ disease in pre-Columbian South America. Am J Phys Anthropol. 68 (1985), pp. 495–498.
J. C. P. Diaz, C. J. Schofield. Emerging Chagas Disease. History of Chagas Disease as a Public Health Problem in Latin America A. Teixeira, M. Vinaud, A. Castro, Edts. Sharjah: Benthan Science Publisher. (2011), pp. 1–9.
L. F. Ferreira, A. M. Jansen, A. Araújo. Chagas disease in prehistory. An Acad Bras Cienc. 83 (2011), pp. 1041–1044.
J. F. Walsh, D. H. Molyneux, M. H. Birley. Deforestation: effects on vector-borne disease. Parasitology. 106 (Suppl) (1993), pp. S55–S75.
R. Briceño-León. Chagas disease and globalization of the Amazon. Cad Saude Publica. 23 (Suppl 1) (2007), pp. S33–S40.
M. Dias Pimenta. Noticias do que he o achaque do bicho, diffiniçam do seu crestameto, subimento corrupçaõ, sinaes, & cura atè, o quinto grao, ou intensaõ delle, suas differenças, & coplicaçoés, com que se ajunta. Lisbon: Miguel Manescal; (1707).
U. G. Meneghelli. M. Dias Pimenta. e a história do megaesôfago e do megacólon chagásicos. (1661-1715) Arg Gastroenterol. 33 (1996), pp. 115–121.
M. A. Miles. The discovery of Chagas disease: progress and prejudice. Infect Dis Clin North Am. 18 (2004), pp. 247–260.
L. G. Ferreira. Erário mineral dividido em doze tratados. Dedicado, e oferecido à puríssima, e sereníssima virgem Nossa Senhora da Conceiçaõ. Lisbon: Miguel Rodrigues; 1735.
J. B. von Spix, C. F. P. von Martius. Reise in Brasilien auf Befehl Sr. Majestät Maximilian Joseph I. Königs von Baiern in den Jahren 1817 bis 1820 gemacht und beschrieben. München: M. Lindauer (1823-1831).
D. P. Kidder, J. Fletcher. Brazil and the Brazilians, Portrayed in Historical and Descriptive Sketches. Philadelphia: Childs & Peterson. (1857).
T. J. H. Langgaard. Diccionario de medicina domestica e popular. Rio de Janeiro: Eduardo & Henrique Laemmert.1865.
F. Guerra. American trypanosomiasis. An historical and a human lesson. Am J Trop Med Hyg. 73 (1970), pp. 83–104.
C. Darwin. Journal of Researches into the Geology and Natural History of the Various Countries Visited by H.M.S. Beagle, under the Command of Captain Fitzroy, R.N. from 1832 to 1836. London: Henry Colburn. (1839).
C. Chagas. Nova tripanozomiaze humana. Estudos sobre a morfolojia e o ciclo evolutivo de Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homem. Mem Inst Oswaldo Cruz. 1 (1909), pp. 159–218.
C. Chagas. Neue Trypanosomen: Vorläufige Mitteilung. Arch Schiffs Trop Hyg. 13 (1909), pp. 120–122.
A. Prata. Evolution of the clinical and epidemiological knowledge about Chagas disease 90 years after its discovery. Mem Inst Oswaldo Cruz. 94(Suppl I) (1999), 81–88.
M. de Lana, C. A. Chiari, E. Chiari, C. M. Morel, A. M. Gonçalves, A. J. Romanha. Characterization of two isolates of Trypanosoma cruzi obtained from the patient Berenice, the first human case of Chagas’ disease described by Carlos Chagas in 1909. Parasitol Res. 82 (1996), pp. 257–260.
C. Chagas. Sobre um tripanossoma do tatu, Tatusia novemcincta, transmitido pela Triatoma geniculata Latr. (1811). possibilidade de ser o tatu um depositário do Trypanosoma cruzi no mundo exterior. Brazil Med. 26 (1811), pp. 305–306.
C. M. Morel. Chagas disease, from discovery to control – and beyond: history, myths and lessons to take home. Mem Inst Oswaldo Cruz. 94 (Suppl I) (1999), pp. 3–16.
S. P. Kropf, M. R. Sá. The discovery of Trypanosoma cruzi and Chagas disease (1908-1909): tropical medicine in Brazil. Hist Cienc Saude Manguinhos. 16 (supl 1): (2009), pp. 13–34.
G. Vianna. Contribuição para o estudo da anatomia patolojica da “Molestia de Carlos Chagas”. Mem Inst Oswaldo Cruz. 3 (1911), pp. 276–294.
E. Brumpt. Le Trypanosoma cruzi évolue chez Conorhinus megistus, Cimex lectularis, Cimex bouetiet Ornithodorus moubata. Cycle évolutif de ce parasite. Bull Soc Pathol Exot. 5 (1912), pp. 360–367.
R. Lewinsohn. Prophet in his own country. Carlos Chagas and the Nobel Perspect Biol Med. 46 (2003), pp. 532–549.
S. Mazza. Casos agudos benignos de enfermedad de Chagas comprobados en la Provincia de Jujuy. MEPRA. 17 (1934), pp. 3–11.
S. Mazza. Nota sobre el primer centenar de formas agudas de enfermedad de Chagas comprobadas en la República por la Misión de Estudios de Patología Regional Argentina. Prensa Med Argent. 1936(23) (1936), pp. 1979–1981.
R. E. Gürtler, L. Diotaiuti, U. Kitron. Commentary: Chagas disease: 100 years since the discovery and lessons for the future. Int J Epidemiol. 37 (2008), pp. 698–701.
J. R. Coura, S. L. de Castro. A critical review on Chagas disease chemotherapy. Mem Inst Oswaldo Cruz. 97 (2002), pp. 3–24.
D. Steverding. The development of drugs for treatment of sleeping sickness: a historical review. Parasit Vectors. 3 (2010), pp. 15.
V. Nussenzweig, R. Sonntag, A. Biancalana, J. Pedreira de Freitas, V. Amato Neto, J. Kloetzel. Ação de corantes tri-fenil-metânicos sobre o Trypanosoma cruziin vitro. Emprego da violeta de genciana na profilaxia da transmissão da moléstia de Chagas por transfusão de sangue. O Hosp. 44 (1953), pp. 731–744.
J. C. P. Dias, C. J. Schofield. The evolution of Chagas disease (American trypanosomiasis) control after 90 years since Carlos Chagas discovery. Mem Inst Oswaldo Cruz. 94 (Suppl I) (1999), pp. 103–121.
J. C. P. Dias, A. C. Silveira, C. J. Schofield. The impact of Chagas disease control in Latin America – a review. Mem Inst Oswaldo Cruz. 97 (2002), pp. 603–612.
P. C. M. Pereira, E. C. Navarro. Challenges and perspectives of Chagas disease: a review. J Venom Anim Toxins Incl Trop Dis. 19 (2013), pp. 34.
K. Hashimoto, C. J. Schofield. Elimination of Rhodnius proxilus in Central America. Parasit Vectors. 5 (2012), pp. 45.
R. de Souza-Lima, M. Barbosa, J. Coura, A. Arcanjo, A. Nascimento, J. Ferreira y col Outbreak of acute Chagas disease associated with oral transmission in the Rio Negro region, Brazilian Amazon. Rev Soc Bras Med Trop. 46 (2013), pp.510–514.
N. Yoshida, K. M. Tyler, M. S. Llewellyn. Invasion mechanisms among emerging food-borne protozoan parasites. Trends Parasitol. 27 (2011), pp. 459–466.
A. Samuels, E. Clark, G. Galdos-C., R. Wiegand, L. Ferrufino, S. Menacho, y col. Epidemiology of and impact of insecticide spraying on Chagas disease in communities in the Bolivian Chaco. PLoS Negl Trop Dis.7 (2013), pp. e2358.
M. Germano, G. Roca-Acevedo, G. Mougabure-C., A. Toloza, C. Vassena, M. Picollo. New findings of insecticide resistance in Triatoma infestans (Heteroptera: Reduviidae) from the Gran Chaco. J Med Entomol. 47 (2010), pp. 1077–1081.
Secretaria de Vigilância em Saúde, Ministério da Saúde. Consenso brasileiro em doença de Chagas. Rev Soc Bras Med Trop. 38 (Supl III): (2005), pp. 7–29.
C. Bern, S. Montgomery, B. Herwaldt, A. Rassi Jr, J. Marin-Neto, R. Dantas, y col, Evaluation and treatment of Chagas disease in the United States: a systematic review. J Am Med Assoc. 98 (2007), pp. 2171–2181.
M. Castillo-Riquelme, F. Guhl, B. Turriago, N. Pinto, F. Rosas, M. Martínez, y col. The costs of preventing and treating Chagas disease in Colombia. PLoS Negl Trop Dis. 2 (2008), pp. e336.
G. A. Schmunis, Z. E. Yadon. Chagas disease: a Latin America health problem becoming a world health problem. Acta Trop. 115 (2010), pp. 14–21.
J. Strasen, T. Williams, G. Ertl, T. Zoller, A. Stich, O. Ritter. Epidemiology of Chagas disease in Europe: many calculations, little knowledge. Clin Res Cardiol. 103 (2014), 1–10.
JA O'Daly. A synthetic media for the growth of Trypanosoma cruzi. Ciba Foundation Symposium on Trypanosomiasis and Leishmaniasis. B. A Newton Ed. (1973), pp. 283.
J. A. O'Daly. A new liquid medium for the growth of Trypanosoma (Schizotrypanum) cruzi (Chagas 1909). Journal of Protozoology 22 (1975), pp. 265 270.
J. A. O'Daly. Serum proteins promoting 3H thymidine uptake by Trypanosoma (Schizotrypanum) cruzi (Chagas) in vitro. Journal of Protozoology 22 (1975), pp. 550 555.
J. A. O'Daly. Effect of fetal calf serum fractions and proteins on division and transformation of T. cruziin vitro. Journal of Protozoology 23 (1976), pp. 577 583.
J. A. O'Daly, A. Bretaña. Ultrastructural observations of Trypanosoma cruzi in a liquid medium: The kinetoplast mitochondrion in division forms. International Journal for Parasitology 6 (1976), pp. 271 278.
A. Bretaña, J. A. O'Daly. Uptake of fetal proteins by Trypanosoma cruzi immuno-fluorescence and ultrastructural studies. Int. J. for Parasitology 6: (1976), pp. 379 386.
J. A. O'Daly, P. Aso. Trypanosoma cruzi, Leishmania donovani and Leishmania mexicana. Extract factor that lyses mammalian cells. Exper Parasitol 47 (1979), 222 231.
J. A. O'Daly. Molecular biology of T. cruzi, L. mexicana and L. donovani. Tropical Disease Research Series: 3. in vitro cultivation of pathogens of Tropical diseases. David S. Rowe Ed. Schwabe & Co. A.G. Basel. Chapter Twenty nine, (1980), pp. 237 243.
J. A. O' Daly, L. Serrano, M. B. Rodríguez. Free amino acid pool and proteolytic enzymes in Trypanosoma cruzi, cultured in vitro. Int J Parasitol 13 (1983), pp. 433 440.
J. A. O'Daly, M. B. Rodríguez, G. Garlin. Trypanosoma cruzi: Growth requirements at different temperatures in fetal bovine serum or peptide supplemented media. Experimental Parasitology. 64 (1987), pp. 78 87.
J. A. O'Daly, M. B. Rodríguez. Protein and nucleotide contamination of bovine liver catalase used in culture medium explains growth of Trypanosoma cruzi. Transactions of the Royal Society of Tropical Medicine and Hygiene. 81 (1987), pp. 1 2.
J. A. O'Daly, S. Simonis, N. De Rolo, H. Caballero. Suppression of humoral immunity and lymphocyte responsiveness during experimental Trypanosoma cruzi infections. Revista do Instituto Medicina Tropical. Sao Paulo, Brasil, 26 (1984), pp. 67 77
L. E. Serrano, J. A. O'Daly. Differential effect of culture epimastigotes and blood form trypomastigotes on normal mouse splenocytes responsiveness to mitogens. Memorias do Instituto Oswaldo Cruz 81 (1986), pp. 207 213.
L. E. Serrano, J. A. O'Daly. Protein fraction from Trypanosoma cruzi infected spleen cell supernatants with immunosuppressive activity in vitro. International Journal for Parasitology. 17 (1987), pp. 851 861.
L. E. Serrano, J. A. O’Daly. Splenocyte membrane changes and immunosupression during infection and reinfection with Trypanosoma cruzi. Investigación Clínica. 31 (1990), pp. 17 31.
J. A. O'Daly, N. Polanco. Variability of Trypanosoma cruzi epimastigote surface antigens with changes in the temperature of the cultures. American Journal of Tropical Medicine and Hygiene. 43 (1990), pp. 44 51.
J. A. O’Daly, P. Garcia, M. B. Rodriguez, T. Ovalles. Leishmania promastigotes secreted/excreted products, characterization and protection of BALB/c mice against virulent parasites by vaccination. The Annals of Infectious Diseases 1 (2012), pp. 1-12.
J. A. O'Daly. A comparison of the molecular biology of Trypanosomas and Leishmaniae and its impact on the development of methods for the diagnosis and vaccination of Leishmaniasis and Chagas disease. Biological Research. 26 (1993), 219-224.
N. Girones, M. Fresno. Etiology of Chagas disease myocarditis: autoimmunity, parasite persistence, or both? Trends Parasitol. (2003), pp. 1919–1922.
A. R. Teixeira, Z. Lacava, J. M. Santana, H. Luna. Insertion of Trypanosoma cruzi DNA in the genome of mammal host cell through infection. Rev Soc Bras Med Trop. (1991), pp. 2455–2458.
A. R. L. Teixeira, M. M. Hecht, M. C. Guimaro, A. O. Sousa, N. Nitz. Chagas’ disease: Parasite Persistence and Autoimmunity. Clin Microbiol Rev. 24 (2011), 592–630.
A. Simoes‐Barbosa, A. M. Barros, N. Nitz, E. R. Argañaraz, A. R. Teixeira. Integration of Trypanosoma cruzi kDNA minicircle sequence in the host genome may be associated with autoimmune serum factors in Chagas disease patients. Mem Inst Oswaldo Cruz. 94 (1999), pp. 249–252.
N. Nitz, C. Gomes, R. de Cássia, M. D'Souza-Ault, F. Moreno, L. Lauria-Pires, y col. Heritable integration of kDNA minicircle sequences from Trypanosoma cruzi into the avian genome: insights into human Chagas disease. Cell. 118 (2004), pp.175-186
C. Suárez, J. J. Puigbo, H. Acquatella, F. R. Tortoledo, V. Ruesta. Similitudes y diferencias morfológicas y fisiopatológicas entre la miocardiopatía chagásica y la miocardiopatía de Takotsubo. Gac Méd Caracas. 124 (2016), pp. 206-224
R. U. Carcavallo, I. Galindez-Giron, J. Jurberg, H. Lent. Atlas of Chagas' disease vectors in the Americas. Rio de Janeiro: Editora Fiocruz, Vol 1 (1997)
[16] A. R. L. Teixeira, N. Nitz, M. C. Guimaro, C. Gomes, C. A. Santos‐Buch. Chagas disease. Postgrad Med J. 82(974), (2006), pp. 788–798.